

Transcriptional regulation and signature patterns revealed by microarray analyses of Streptococcus pneumoniae R6 challenged with sublethal concentrations of translation inhibitors
- PMID: 12486074
- PMCID: PMC141824
- DOI: 10.1128/JB.185.1.359-370.2003
Transcriptional regulation and signature patterns revealed by microarray analyses of Streptococcus pneumoniae R6 challenged with sublethal concentrations of translation inhibitors
Abstract
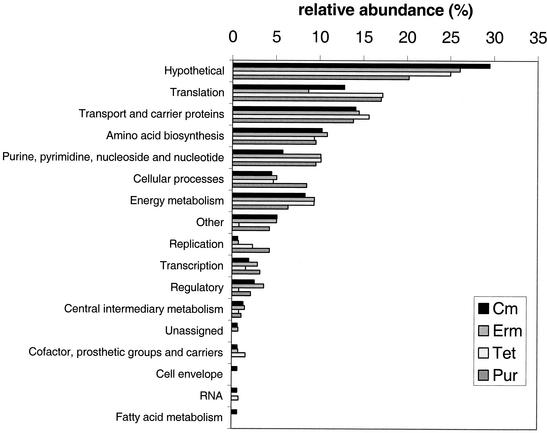
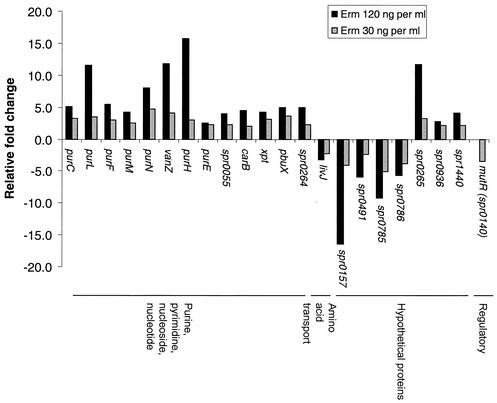
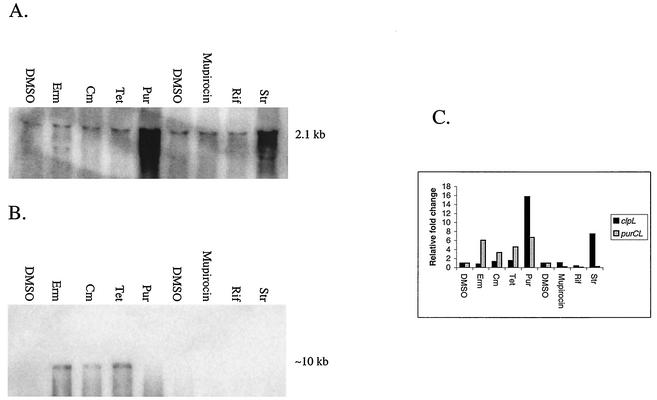
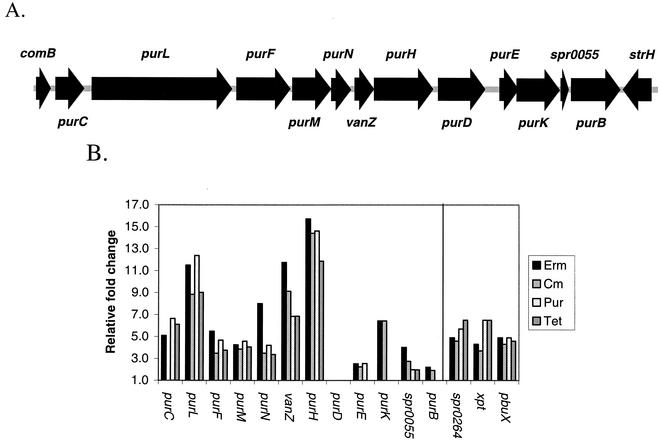
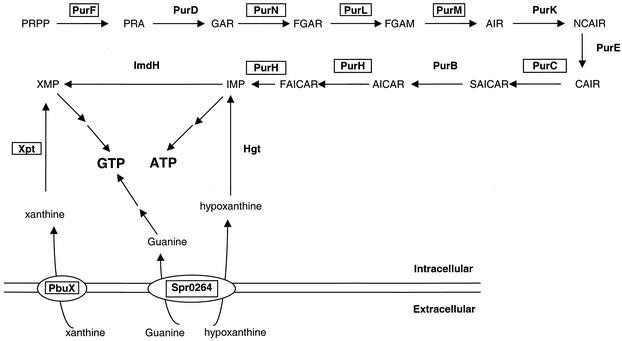
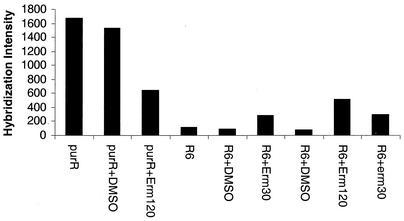
The effects of sublethal concentrations of four different classes of translation inhibitors (puromycin, tetracycline, chloramphenicol, and erythromycin) on global transcription patterns of Streptococcus pneumoniae R6 were determined by microarray analyses. Consistent with the general mode of action of these inhibitors, relative transcript levels of genes that encode ribosomal proteins and translation factors or that mediate tRNA charging and amino acid biosynthesis increased or decreased, respectively. Transcription of the heat shock regulon was induced only by puromycin or streptomycin treatment, which lead to truncation or mistranslation, respectively, but not by other antibiotics that block translation, transcription, or amino acid charging of tRNA. In contrast, relative transcript amounts of certain genes involved in transport, cellular processes, energy metabolism, and purine nucleotide (pur) biosynthesis were changed by different translation inhibitors. In particular, transcript amounts from a pur gene cluster and from purine uptake and salvage genes were significantly elevated by several translation inhibitors, but not by antibiotics that target other cellular processes. Northern blotting confirmed increased transcript amounts from part of the pur gene cluster in cells challenged by translation inhibitors and revealed the presence of a 10-kb transcript. Purine metabolism genes were negatively regulated by a homologue of the PurR regulatory protein, and full derepression in a DeltapurR mutant depended on optimal translation. Unexpectedly, hierarchical clustering of the microarray data distinguished among the global transcription patterns caused by antibiotics that inhibit different steps in the translation cycle. Together, these results show that there is extensive control of transcript amounts by translation in S. pneumoniae, especially for de novo purine nucleotide biosynthesis. In addition, these global transcription patterns form a signature that can be used to classify the mode of action and potential mechanism of new translation inhibitors.
Figures

References
-
- Babitzke, P. 1997. Regulation of tryptophan biosynthesis: Trp-ing the TRAP or how Bacillus subtilis reinvented the wheel. Mol. Microbiol. 26:1-9. - PubMed
-
- Brodersen, D. E., W. M. Clemons, Jr., A. P. Carter, R. J. Morgan-Warren, B. T. Wimberly, and V. Ramakrishnan. 2000. The structural basis for the action of the antibiotics tetracycline, pactamycin, and hygromycin B on the 30S ribosomal subunit. Cell 103:1143-1154. - PubMed
-
- Campbell, E. A., N. Korzheva, A. Mustaev, K. Murakami, S. Nair, A. Goldfarb, and S. A. Darst. 2001. Structural mechanism for rifampicin inhibition of bacterial RNA polymerase. Cell 104:901-912. - PubMed
-
- Carter, A. P., W. M. Clemons, D. E. Brodersen, R. J. Morgan-Warren, B. T. Wimberly, and V. Ramakrishnan. 2000. Functional insights from the structure of the 30S ribosomal subunit and its interactions with antibiotics. Nature 407:340-348. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
