

Sensitization of regulated exocytosis by protein kinase C
- PMID: 12486252
- PMCID: PMC139268
- DOI: 10.1073/pnas.232588899
Sensitization of regulated exocytosis by protein kinase C
Abstract
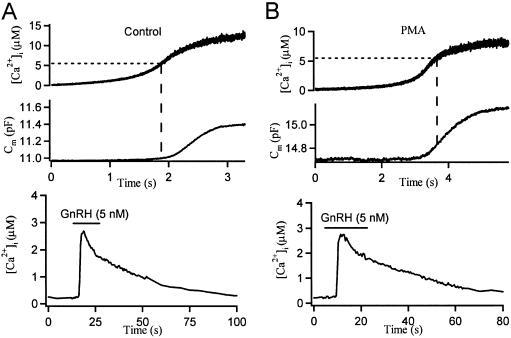
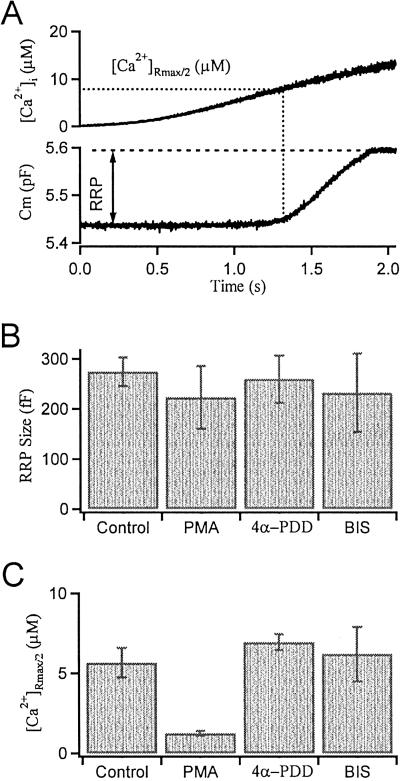
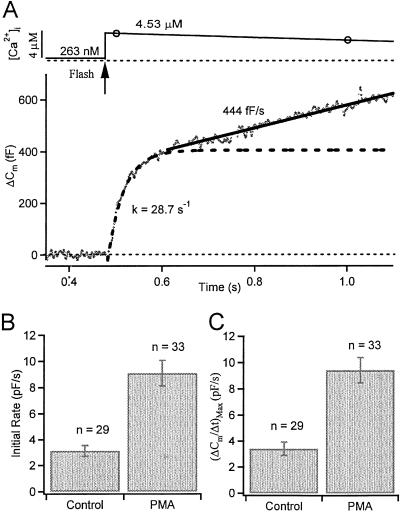
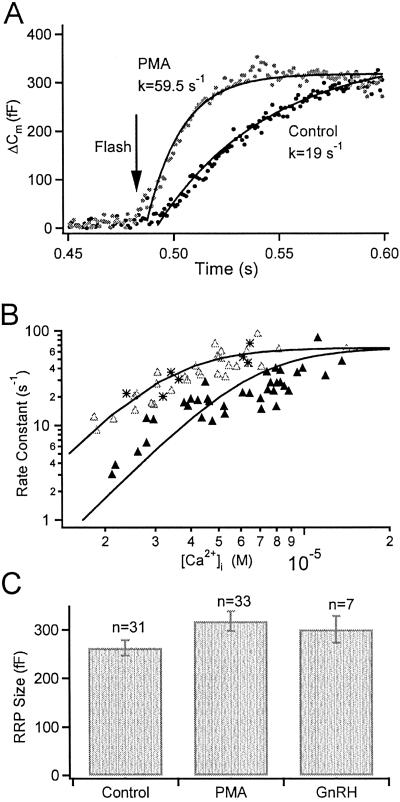
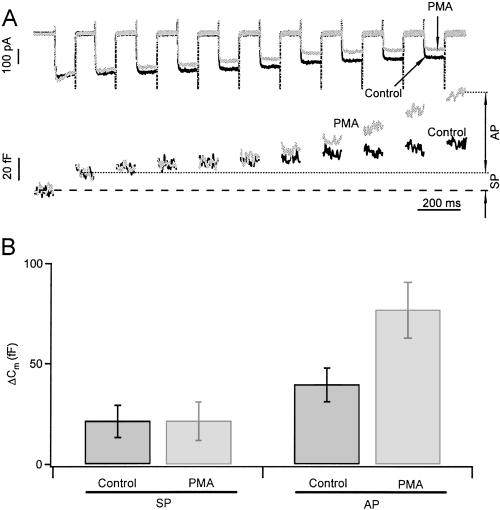
Activation of protein kinase C (PKC) increases vesicular secretion in many cell types. We determined the calcium dependence of secretion and the size of the readily releasable pool of secretory granules in pituitary gonadotropes by photorelease of caged-calcium. The calcium affinity for exocytosis was roughly doubled by activation of PKC by a phorbol ester, whereas the size of the readily releasable pool was not greatly increased. The effect was due to activation of PKC, because it was blocked by a PKC inhibitor and was not mimicked by an inactive phorbol ester analogue. A similar increase in calcium sensitivity was induced by preincubation with gonadotropin-releasing hormone, the physiological releasing hormone. These findings provide direct evidence for physiological regulation of secretion by enhancement of Ca2+-sensing steps. Because exocytosis depends on the third- to fourth-power of intracellular free Ca2+ concentration, this mechanism ensures a powerful up-regulation of hormone release and may explain how PKC can stimulate exocytosis without an increase of Ca2+ above the resting level.
Figures
Comment in
-
Specificity emerges in the dissection of diacylglycerol- and protein kinase C-mediated signalling pathways.Proc Natl Acad Sci U S A. 2002 Dec 24;99(26):16522-3. doi: 10.1073/pnas.022708199. Epub 2002 Dec 16. Proc Natl Acad Sci U S A. 2002. PMID: 12486246 Free PMC article. No abstract available.
References
-
- Chen Y. A., Duvvuri, V., Schulman, H. & Scheller, R. H. (1999) J. Biol. Chem. 274, 26469-26476. - PubMed
-
- Evans G. J. O., Wilkinson, M. C., Graham, M. E., Turner, K. M., Chamberlain, L. H., Burgoyne, R. D. & Morgan, A. (2001) J. Biol. Chem. 276, 47877-47885. - PubMed
-
- Gillis K. D., Mossner, R. & Neher, E. (1996) Neuron 16, 1209-1220. - PubMed
-
- Smith C., Moser, T., Xu, T. & Neher, E. (1998) Neuron 20, 1243-1253. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
