

Entry of the lymphogranuloma venereum strain of Chlamydia trachomatis into host cells involves cholesterol-rich membrane domains
- PMID: 12496174
- PMCID: PMC143347
- DOI: 10.1128/IAI.71.1.260-266.2003
Entry of the lymphogranuloma venereum strain of Chlamydia trachomatis into host cells involves cholesterol-rich membrane domains
Abstract
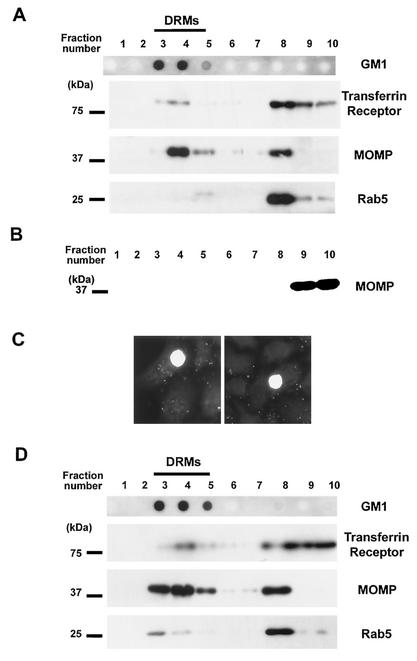
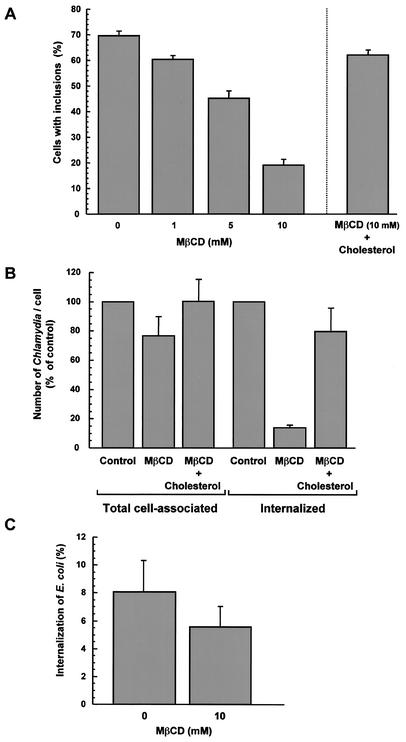
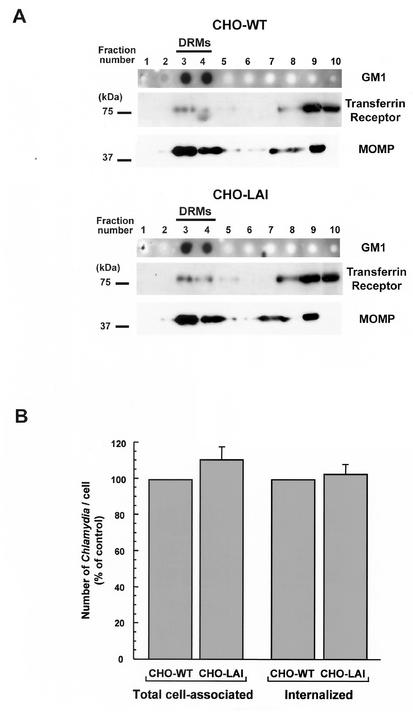
Chlamydiae are bacterial pathogens which develop strictly inside the epithelial cells of their hosts. The mechanism used by chlamydiae to enter cells is not well characterized; however, it is thought to consist of a receptor-mediated process. In addition, the formation of clathrin-coated pits appears to be dispensable for chlamydiae to be internalized by host cells. Clathrin-independent endocytosis has recently been shown to occur through cholesterol-rich lipid microdomains, which are characterized by detergent insolubility. In the present study, we investigated whether these lipid domains play a role in Chlamydia trachomatis serovar L2 internalization by host cells. Our results show that after binding to HeLa cells, chlamydiae are associated with detergent-resistant lipid microdomains (DRMs), which can be isolated by fractionation of infected HeLa cells and flotation on a sucrose gradient. After internalization by HeLa cells, chlamydiae were still found in DRMs. In addition, extraction of plasma membrane cholesterol inhibited infection of HeLa cells by C. trachomatis. Many of the proteins associated with DRMs are glycosylphosphatidylinositol (GPI)-anchored proteins; however, our results could not identify a role for GPI-anchored proteins in the entry process. The same results were obtained for Chlamydia psittaci strain GPIC. We propose that cholesterol-rich domains participate in the entry of chlamydiae into host cells. Chlamydia binding to cholesterol-rich domains may lead to coalescence of the bacterial cells, which could trigger internalization by host cells.
Figures
References
-
- Abrami, L., M. Fivaz, T. Kobayashi, T. Kinoshita, R. G. Parton, and F. G. van der Goot. 2001. Cross-talk between caveolae and glycosylphosphatidylinositol-rich domains. J. Biol. Chem. 276:30729-30736. - PubMed
-
- Baorto, D. M., Z. Gao, R. Malaviya, M. L. Dustin, A. van der Merwe, D. M. Lublin, and S. N. Abraham. 1997. Survival of FimH-expressing enterobacteria in macrophages relies on glycolipid traffic. Nature 389:636-639. - PubMed
-
- Boleti, H., A. Benmerah, D. M. Ojcius, N. Cerf-Bensussan, and A. Dautry-Varsat. 1999. Chlamydia infection of epithelial cells expressing dynamin and Eps15 mutants: clathrin-independent entry into cells and dynamin-dependent productive growth. J. Cell Sci. 112:1487-1496. - PubMed
-
- Brown, D. A., and E. London. 2000. Structure and function of sphingolipid- and cholesterol-rich membrane rafts. J. Biol. Chem. 275:17221-17224. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
