

LIN-39/Hox triggers cell division and represses EFF-1/fusogen-dependent vulval cell fusion
- PMID: 12502736
- PMCID: PMC187502
- DOI: 10.1101/gad.251202
LIN-39/Hox triggers cell division and represses EFF-1/fusogen-dependent vulval cell fusion
Abstract
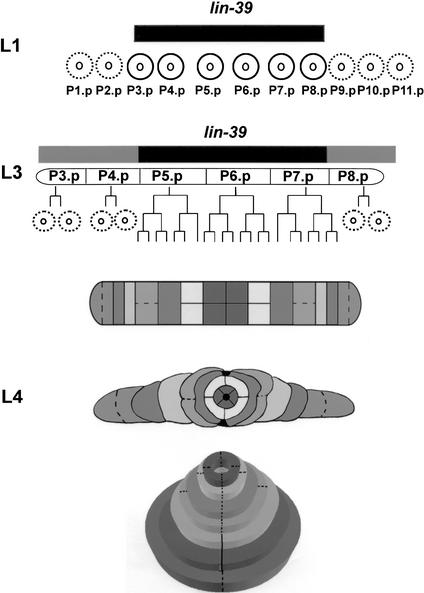
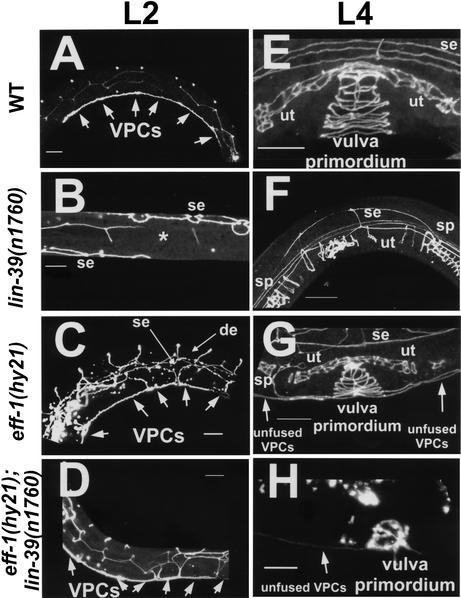
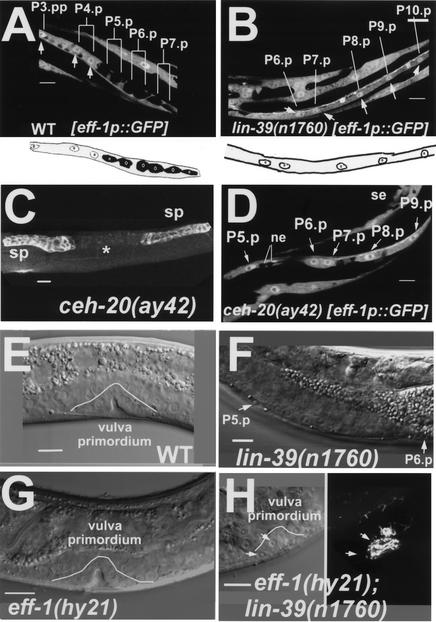
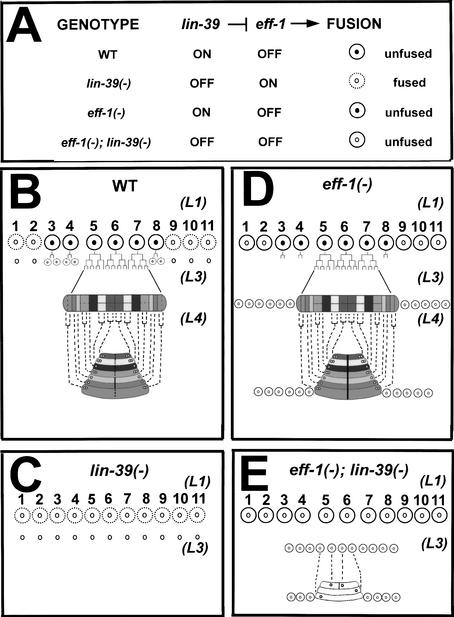
General mechanisms by which Hox genes establish cell fates are known. However, a few Hox effectors mediating cell behaviors have been identified. Here we found the first effector of LIN-39/HoxD4/Dfd in Caenorhabditis elegans. In specific vulval precursor cells (VPCs), LIN-39 represses early and late expression of EFF-1, a membrane protein essential for cell fusion. Repression of eff-1 is also achieved by the activity of CEH-20/Exd/Pbx, a known cofactor of Hox proteins. Unfused VPCs in lin-39(-);eff-1(-) double mutants fail to divide but migrate, executing vulval fates. Thus, lin-39 is essential for inhibition of EFF-1-dependent cell fusion and stimulation of cell proliferation during vulva formation. Supplemental material is available at http://www.genesdev.org.
Figures
References
-
- Alper S, Kenyon C. REF-1, a protein with two bHLH domains, alters the pattern of cell fusion in C. elegans by regulating Hox protein activity. Development. 2001;128:1793–1804. - PubMed
-
- ————— The zinc finger protein REF-2 functions with the Hox genes to inhibit cell fusion in the ventral epidermis of C. elegans. Development. 2002;129:3335–3348. - PubMed
-
- Amthor H, Christ B, Weil M, Patel K. The importance of timing differentiation during limb muscle development. Curr Biol. 1998;8:642–652. - PubMed
-
- Ch'ng Q, Kenyon C. egl-27 generates anteroposterior patterns of cell fusion in C. elegans by regulating Hox gene expression and Hox protein function. Development. 1999;126:3303–3312. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources