

A macromolecular complex of beta 2 adrenergic receptor, CFTR, and ezrin/radixin/moesin-binding phosphoprotein 50 is regulated by PKA
- PMID: 12502786
- PMCID: PMC140971
- DOI: 10.1073/pnas.0135434100
A macromolecular complex of beta 2 adrenergic receptor, CFTR, and ezrin/radixin/moesin-binding phosphoprotein 50 is regulated by PKA
Abstract
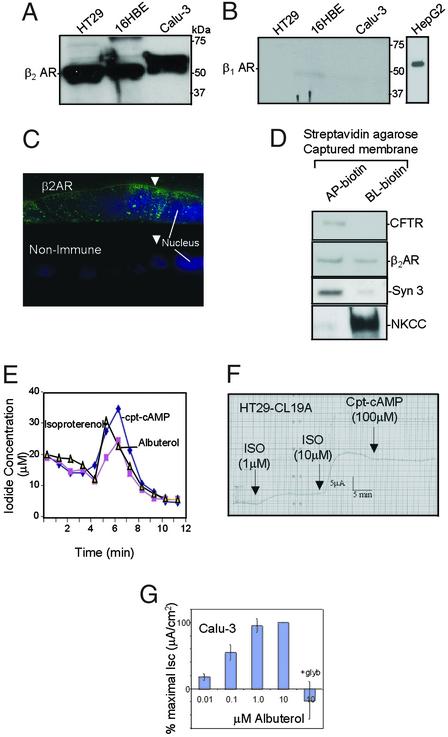
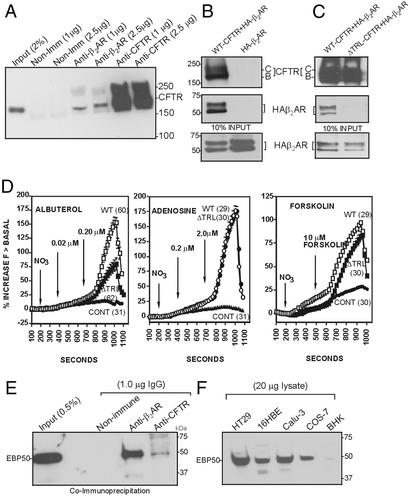
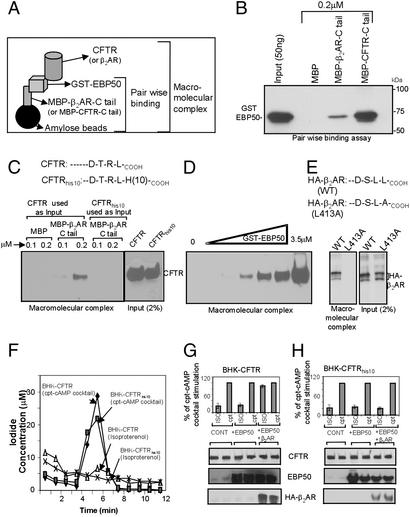
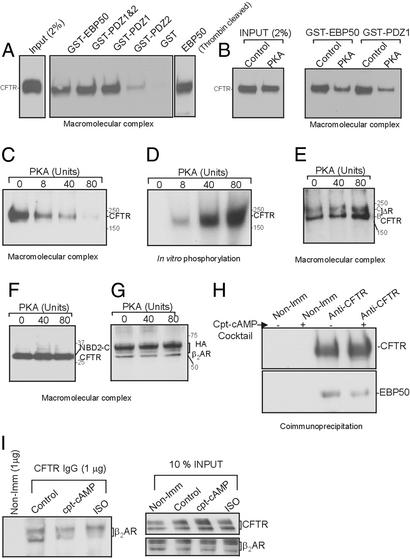
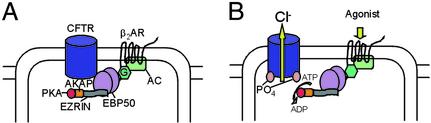
It has been demonstrated previously that both the cystic fibrosis transmembrane conductance regulator (CFTR) and beta(2) adrenergic receptor (beta(2)AR) can bind ezrinradixinmoesin-binding phosphoprotein 50 (EBP50, also referred to as NHERF) through their PDZ motifs. Here, we show that beta(2) is the major adrenergic receptor isoform expressed in airway epithelia and that it colocalizes with CFTR at the apical membrane. beta(2)AR stimulation increases CFTR activity, in airway epithelial cells, that is glybenclamide sensitive. Deletion of the PDZ motif from CFTR uncouples the channel from the receptor both physically and functionally. This uncoupling is specific to the beta(2)AR receptor and does not affect CFTR coupling to other receptors (e.g., adenosine receptor pathway). Biochemical studies demonstrate the existence of a macromolecular complex involving CFTR-EBP50-beta(2)AR through PDZ-based interactions. Assembly of the complex is regulated by PKA-dependent phosphorylation. Deleting the regulatory domain of CFTR abolishes PKA regulation of complex assembly. This report summarizes a macromolecular signaling complex involving CFTR, the implications of which may be relevant to CFTR-dysfunction diseases.
Figures
References
-
- Welsh M J, Tsui L-C, Boat T F, Beaudet A L. In: The Metabolic and Molecular Basis of Inherited Diseases: Membrane Transport Systems. Scriver C, Beaudet A L, Sly W S, Valle D, editors. Vol. 3. New York: McGraw–Hill; 1995. pp. 3799–3876.
-
- Gadsby D C, Nairn A C. Adv Second Messenger Phosphoprotein Res. 1999;33:79–106. - PubMed
-
- Kunzelmann K. News Physiol Sci. 2001;16:167–170. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
