

Mitotic clonal expansion: a synchronous process required for adipogenesis
- PMID: 12502791
- PMCID: PMC140878
- DOI: 10.1073/pnas.0137044100
Mitotic clonal expansion: a synchronous process required for adipogenesis
Abstract
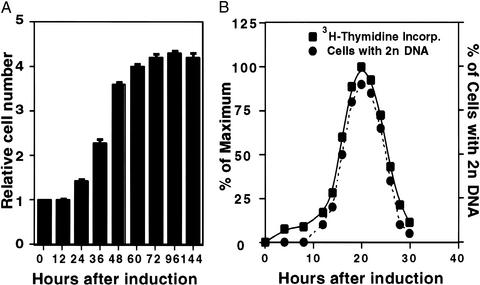
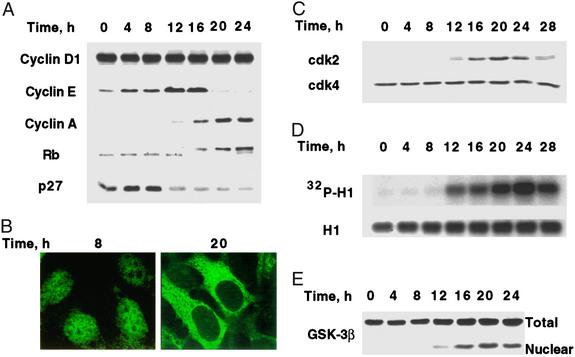
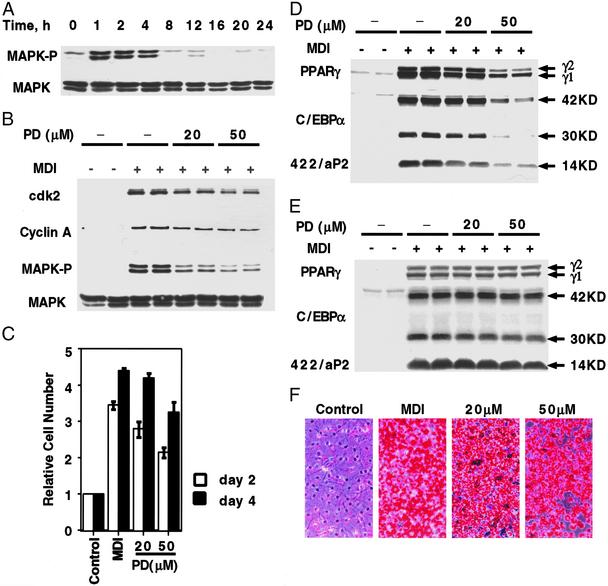
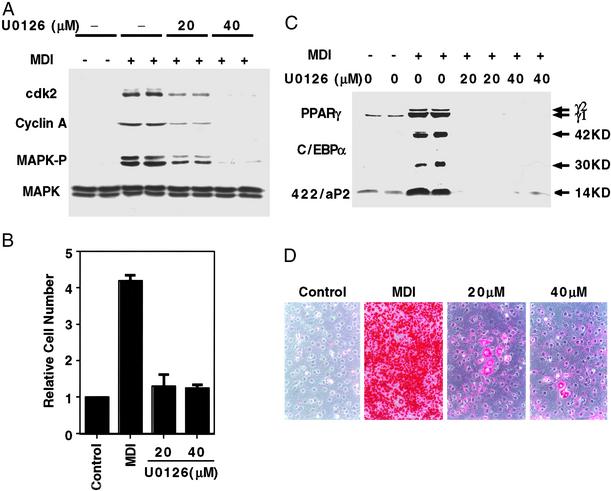
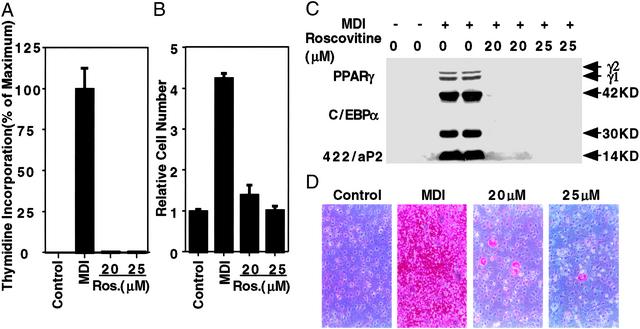
When induced to differentiate, growth-arrested 3T3-L1 preadipocytes synchronously reenter the cell cycle and undergo mitotic clonal expansion (MCE) followed by expression of genes that produce the adipocyte phenotype. The preadipocytes traverse the G(1)S checkpoint synchronously as evidenced by the expressionactivation of cdk2-cyclin-EA, turnover of p27kip1, hyperphosphorylation of Rb, translocation of cyclin D(1) from nuclei to cytoplasm and GSK-3beta from cytoplasm to nuclei, and incorporation of [(3)H]thymidine into DNA. As the cells cross the G(1)S checkpoint, CEBPbeta acquires DNA-binding activity, initiating a cascade of transcriptional activation that culminates in the expression of adipocyte proteins. The mitogen-activated protein kinaseextracellular signal-regulated kinase kinase (MEK) inhibitor PD98059 delays, but does not block, MCE and differentiation, the extent of the delay causing a comparable delay in the expression of cell-cycle markers, MCE, and adipogenesis. The more potent and specific MEK inhibitor UO126 and the cyclin-dependent kinase inhibitor roscovitine, which inhibit the cell cycle at different points, block MCE, expression of cell cycle and adipocyte markers, as well as adipogenesis. These results show that MCE is a prerequisite for differentiation of 3T3-L1 preadipocytes into adipocytes.
Figures
References
-
- Shepherd P R, Gnudi L, Tozzo E, Yang H, Leach F, Kahn B B. J Biol Chem. 1993;268:22243–22246. - PubMed
-
- Gnudi L, Shepherd P R, Kahn B B. Proc Nutr Soc. 1996;55:191–199. - PubMed
-
- Bernlohr D A, Bolanowski M A, Kelly T J, Lane M D. J Biol Chem. 1985;260:5563–5567. - PubMed
-
- Cornelius P, MacDougald O A, Lane M D. Annu Rev Nutr. 1994;14:99–129. - PubMed
-
- MacDougald O A, Lane M D. Annu Rev Biochem. 1995;64:345–373. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
