

Control of protein life-span by N-terminal methionine excision
- PMID: 12505980
- PMCID: PMC140049
- DOI: 10.1093/emboj/cdg007
Control of protein life-span by N-terminal methionine excision
Abstract
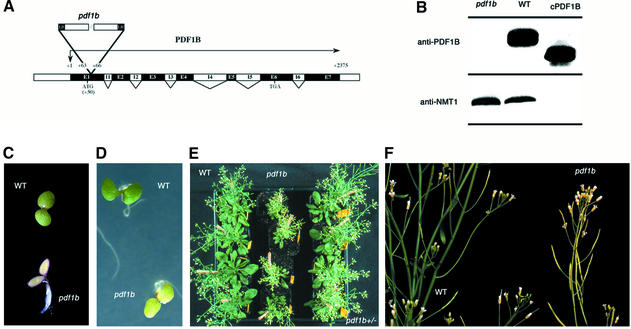
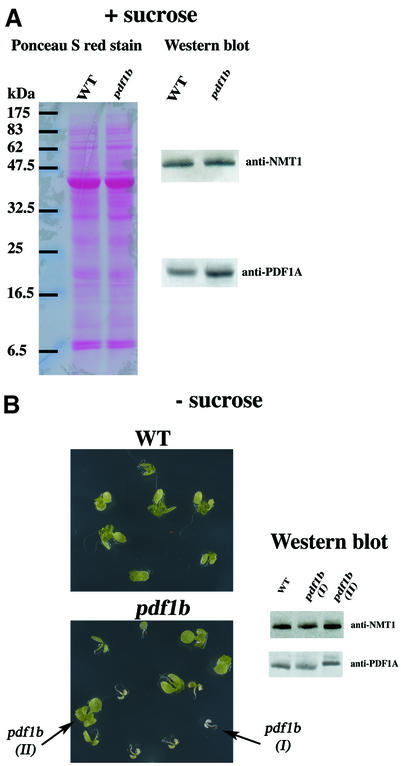
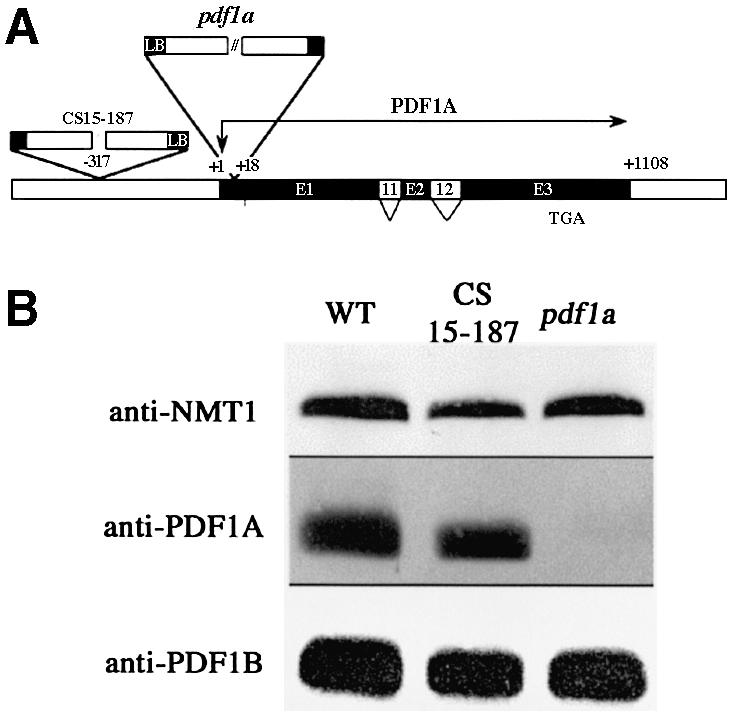
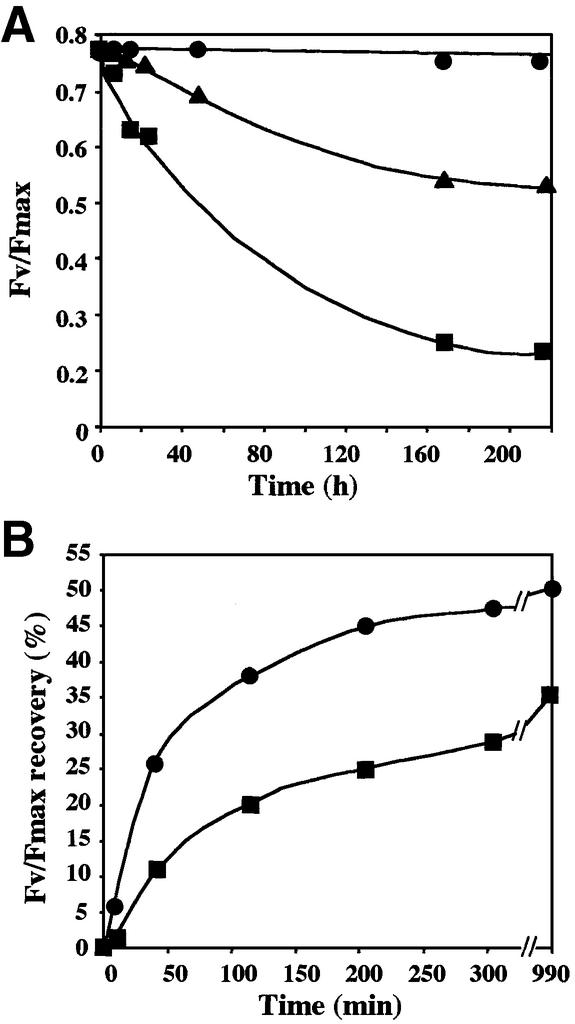
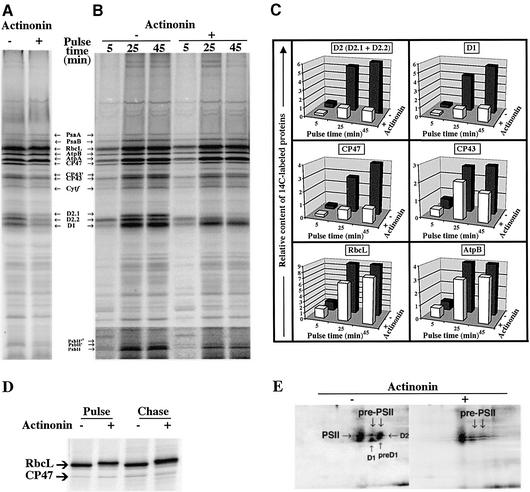
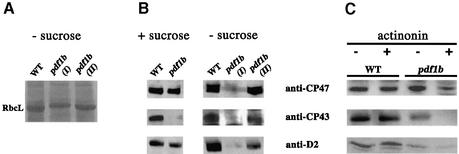
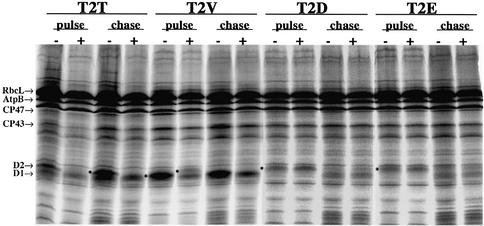
Peptide deformylases (PDFs) have been discovered recently in eukaryotic genomes, and it appears that N-terminal methionine excision (NME) is a conserved pathway in all compartments where protein synthesis occurs. This work aimed at uncovering the function(s) of NME in a whole proteome, using the chloroplast-encoded proteins of both Arabidopsis thaliana and Chlamydomonas reinhardtii as model systems. Disruption of PDF1B in A.thaliana led to an albino phenotype, and an extreme sensitivity to the PDF- specific inhibitor actinonin. In contrast, a knockout line for PDF1A exhibited no apparent phenotype. Photosystem II activity in C.reinhardtii cells was substantially reduced by the presence of actinonin. Pulse-chase experiments revealed that PDF inhibition leads to destabilization of a crucial subset of chloroplast-encoded photosystem II components in C. reinhardtii. The same proteins were destabilized in pdf1b. Site-directed substitutions altering NME of the most sensitive target, subunit D2, resulted in similar effects. Thus, plastid NME is a critical mechanism specifically influencing the life-span of photosystem II polypeptides. A general role of NME in modulating the half-life of key subsets of proteins is suggested.
Figures

References
-
- Adam Z. and Clarke,A.K. (2002) Cutting edge of chloroplast proteolysis. Trends Plant Sci., 7, 451–456. - PubMed
-
- Aro E.M., Virgin,I. and Andersson,B. (1993) Photoinhibition of photosystem II. Inactivation, protein damage and turnover. Biochim. Biophys. Acta, 1143, 113–134. - PubMed
-
- Biran D., Gur,E., Gollan,L. and Ron,E.Z. (2000) Control of methionine biosynthesis in Escherichia coli by proteolysis. Mol. Microbiol., 37, 1436–1443. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
