

TRPM7 provides an ion channel mechanism for cellular entry of trace metal ions
- PMID: 12508053
- PMCID: PMC2217320
- DOI: 10.1085/jgp.20028740
TRPM7 provides an ion channel mechanism for cellular entry of trace metal ions
Abstract
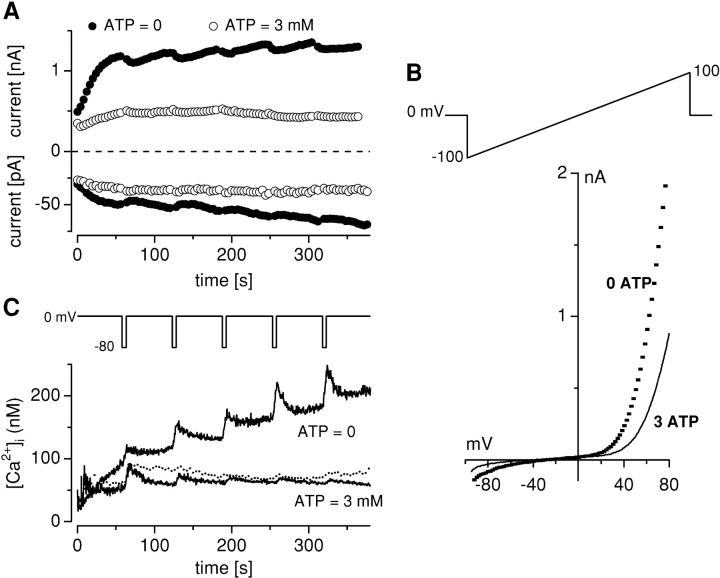
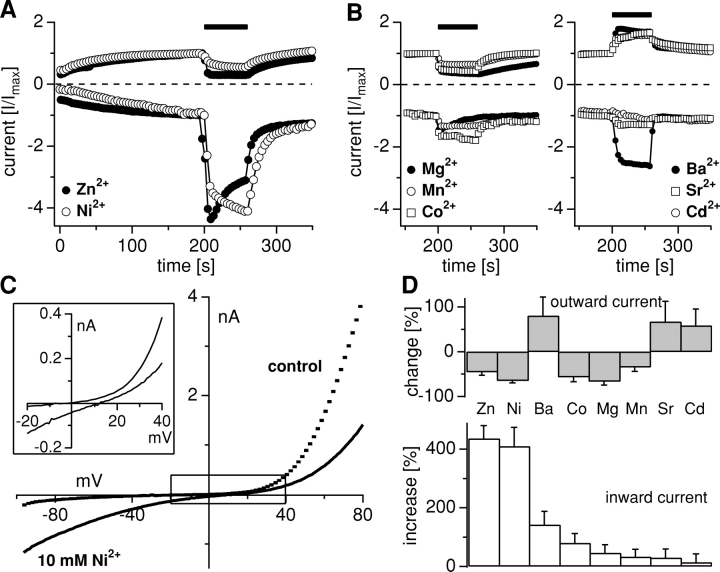
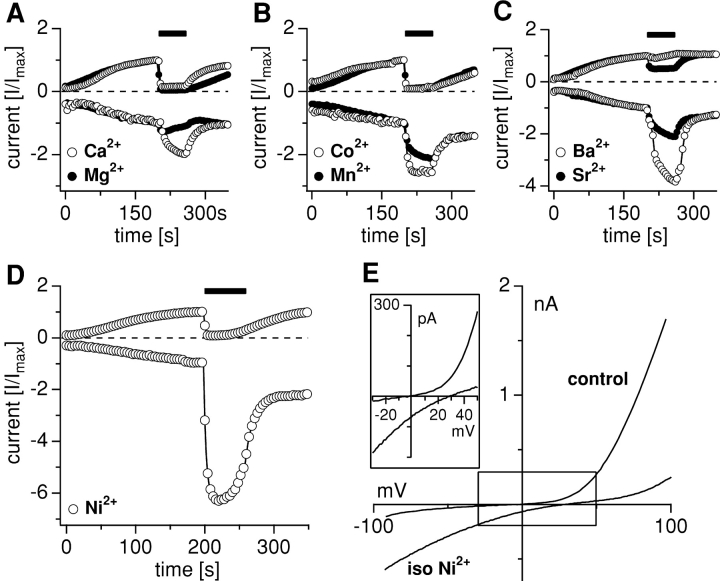
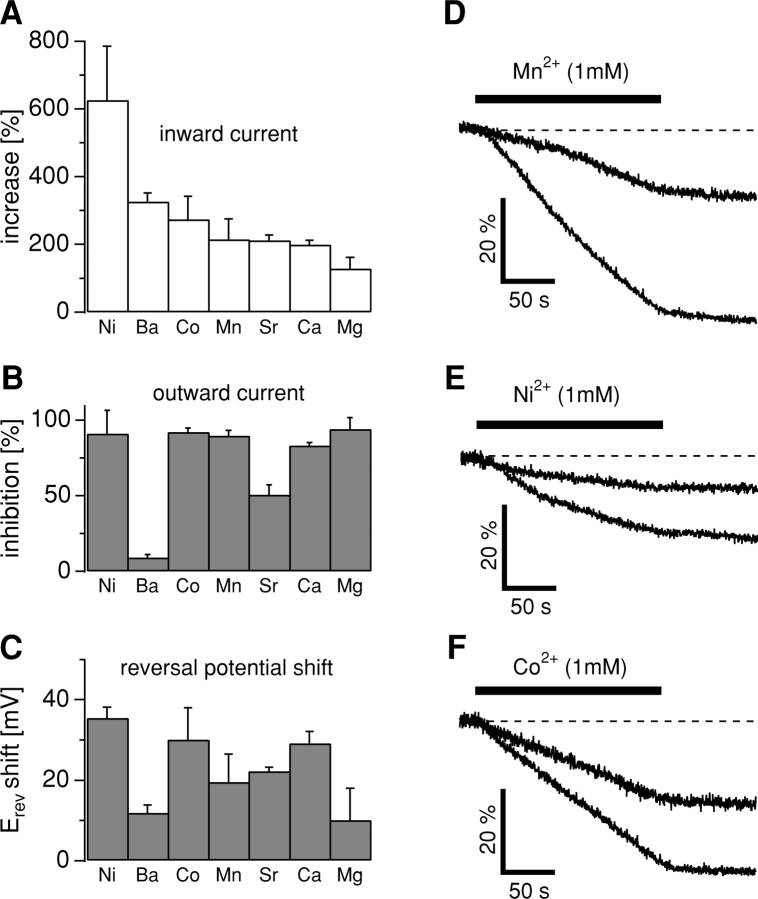
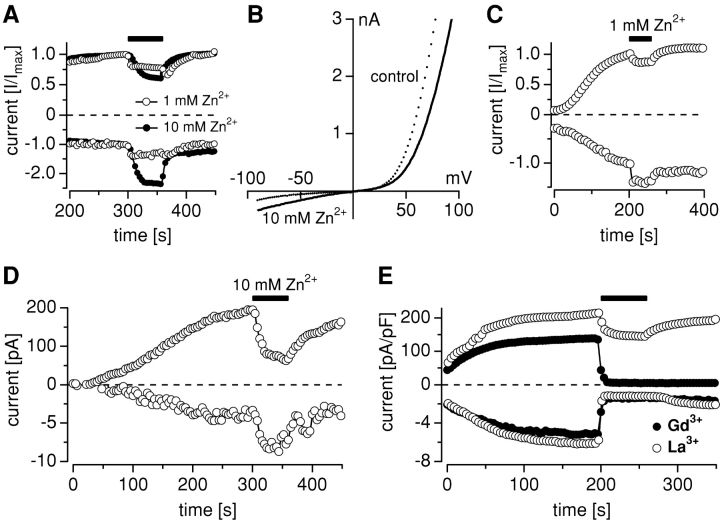
Trace metal ions such as Zn(2+), Fe(2+), Cu(2+), Mn(2+), and Co(2+) are required cofactors for many essential cellular enzymes, yet little is known about the mechanisms through which they enter into cells. We have shown previously that the widely expressed ion channel TRPM7 (LTRPC7, ChaK1, TRP-PLIK) functions as a Ca(2+)- and Mg(2+)-permeable cation channel, whose activity is regulated by intracellular Mg(2+) and Mg(2+).ATP and have designated native TRPM7-mediated currents as magnesium-nucleotide-regulated metal ion currents (MagNuM). Here we report that heterologously overexpressed TRPM7 in HEK-293 cells conducts a range of essential and toxic divalent metal ions with strong preference for Zn(2+) and Ni(2+), which both permeate TRPM7 up to four times better than Ca(2+). Similarly, native MagNuM currents are also able to support Zn(2+) entry. Furthermore, TRPM7 allows other essential metals such as Mn(2+) and Co(2+) to permeate, and permits significant entry of nonphysiologic or toxic metals such as Cd(2+), Ba(2+), and Sr(2+). Equimolar replacement studies substituting 10 mM Ca(2+) with the respective divalent ions reveal a unique permeation profile for TRPM7 with a permeability sequence of Zn(2+) approximately Ni(2+) >> Ba(2+) > Co(2+) > Mg(2+) >/= Mn(2+) >/= Sr(2+) >/= Cd(2+) >/= Ca(2+), while trivalent ions such as La(3+) and Gd(3+) are not measurably permeable. With the exception of Mg(2+), which exerts strong negative feedback from the intracellular side of the pore, this sequence is faithfully maintained when isotonic solutions of these divalent cations are used. Fura-2 quenching experiments with Mn(2+), Co(2+), or Ni(2+) suggest that these can be transported by TRPM7 in the presence of physiological levels of Ca(2+) and Mg(2+), suggesting that TRPM7 represents a novel ion-channel mechanism for cellular metal ion entry into vertebrate cells.
Figures
References
-
- Berridge, M., P. Lipp, and M. Bootman. 1999. Calcium signalling. Curr. Biol. 9:R157–R159. - PubMed
-
- Caterina, M.J., M.A. Schumacher, M. Tominaga, T.A. Rosen, J.D. Levine, and D. Julius. 1997. The capsaicin receptor: a heat-activated ion channel in the pain pathway. Nature. 389:816–824. - PubMed
-
- Choi, D.W., and J.Y. Koh. 1998. Zinc and brain injury. Annu. Rev. Neurosci. 21:347–375. - PubMed
-
- Cousins, R.J., and R.J. McMahon. 2000. Integrative aspects of zinc transporters. J. Nutr. 130:1384S–1387S. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
