

Functional electron microscopy in studies of plant response and adaptation to anaerobic stress
- PMID: 12509337
- PMCID: PMC4244998
- DOI: 10.1093/aob/mcf244
Functional electron microscopy in studies of plant response and adaptation to anaerobic stress
Abstract
This article reviews the contribution made by functional electron microscopy towards identifying and understanding the reactions of plant roots and shoots to anaerobic stress. Topics examined include: (1) unexpected hypersensitivity, rather than hyper-resistance, to anoxia of root tips of flooding-tolerant plants; (2) protective, rather than damaging, effects of a stimulated energy metabolism (glycolysis and fermentation) under anaerobic conditions; (3) the concept of two main strategies of plant adaptation to anaerobic environments, namely avoidance of anaerobiosis on the whole plant level, termed 'apparent' tolerance, and metabolic adaptation at the cellular and molecular levels, termed 'true' tolerance; (4) the importance of protein synthesis during hypoxia and anoxia for enhanced energy production and metabolic adaptation; (5) a general adaptive syndrome in plants to stress at the ultrastructural level and a possible molecular mechanism for its realization under anoxia; (6) the physiological role of anaerobically synthesized lipids and nitrate as alternative electron acceptors in an oxygen-free medium; and (7) the selection of cell lines derived from callus cultures that possess enhanced tolerance to anoxia and can regenerate whole plants with improved tolerance of soil waterlogging.
Figures
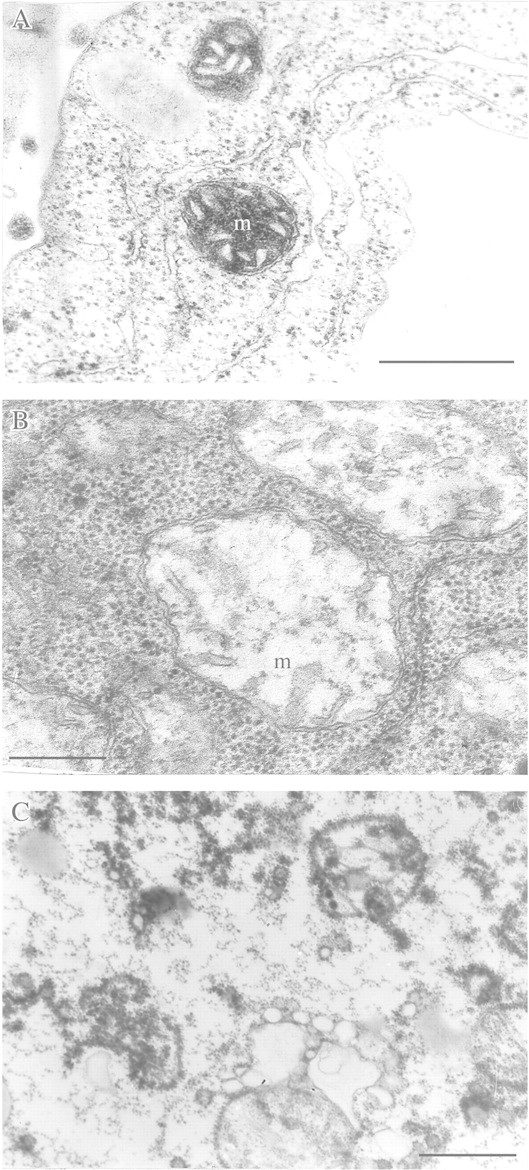
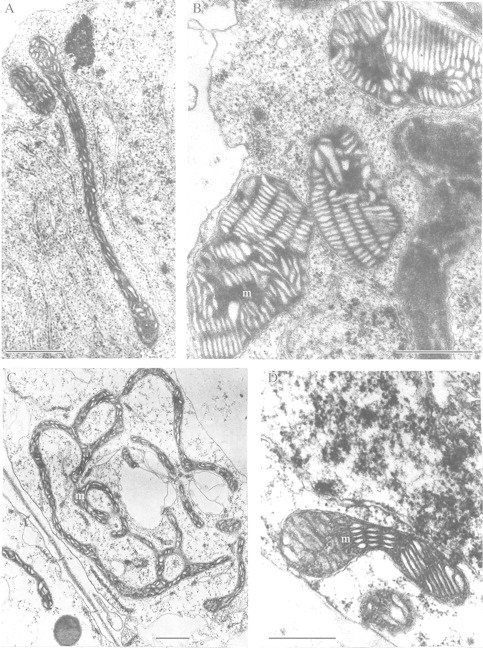
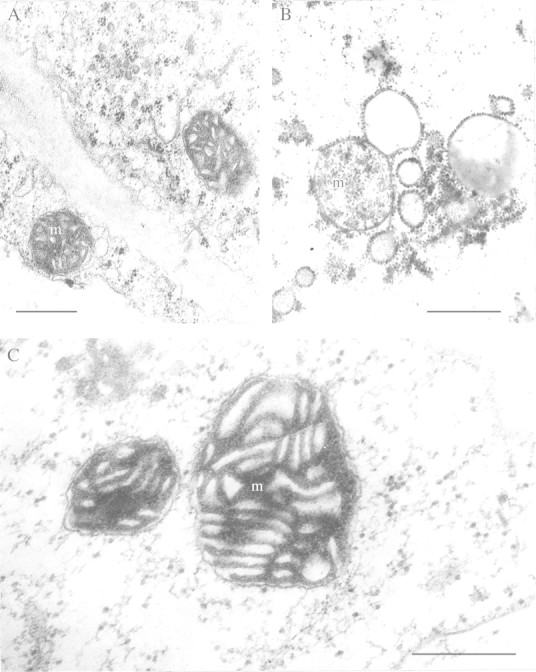
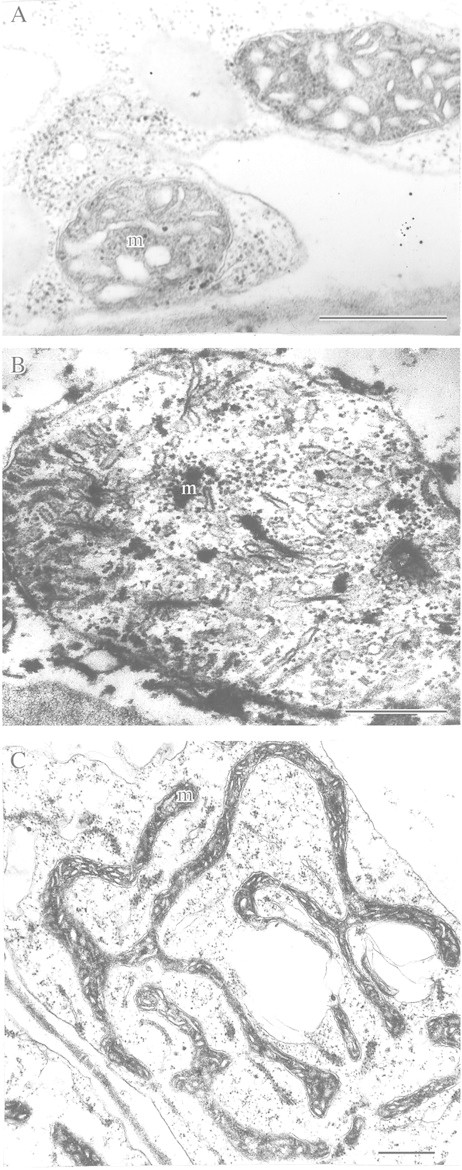
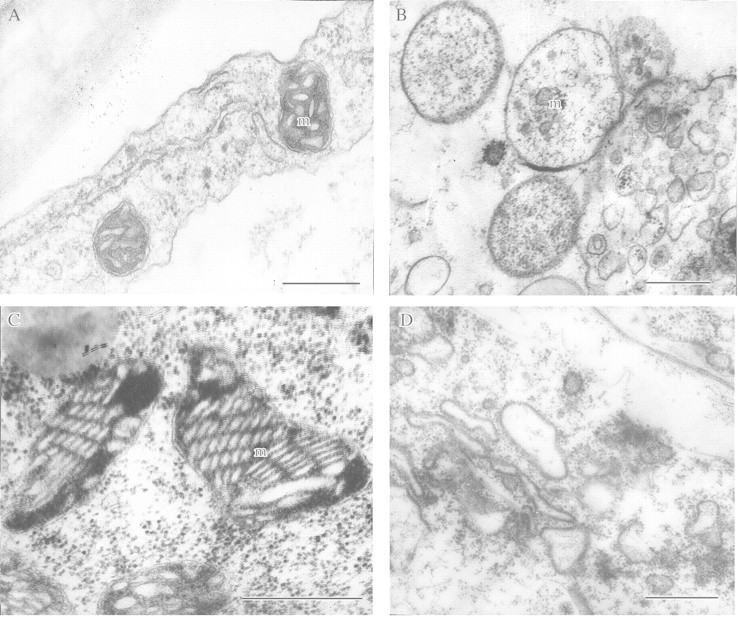
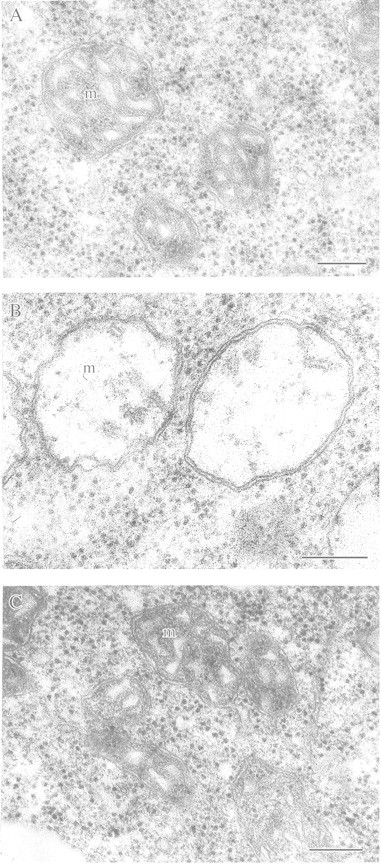
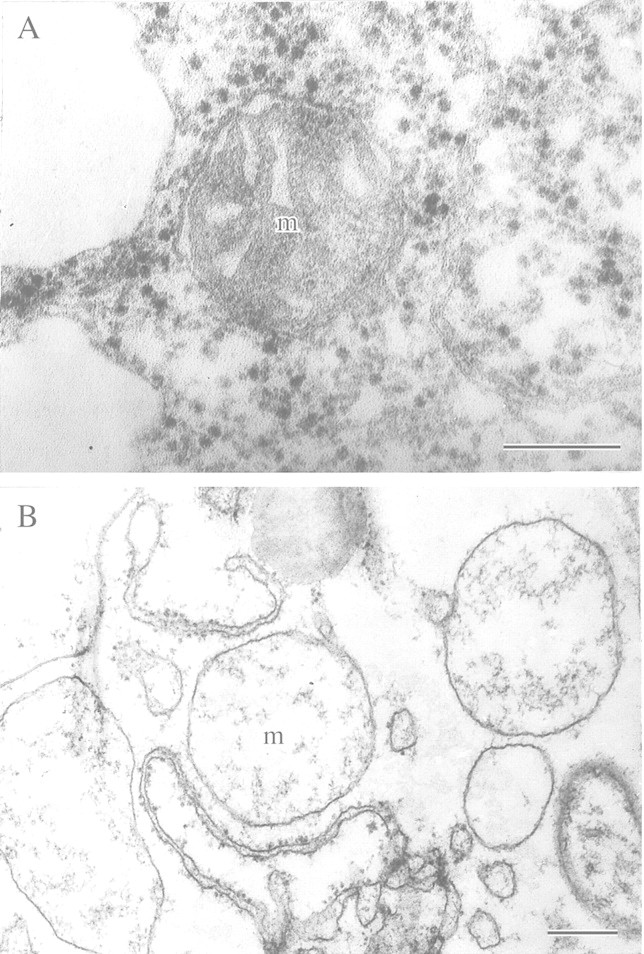
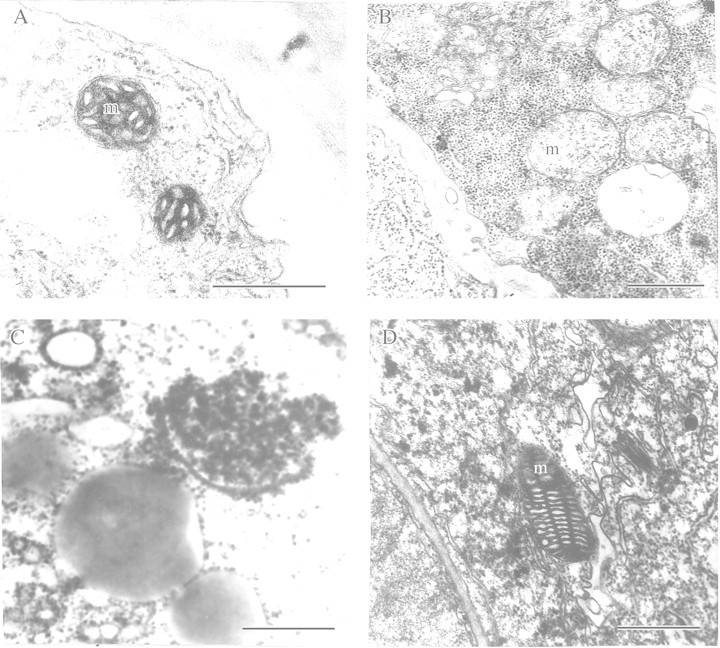
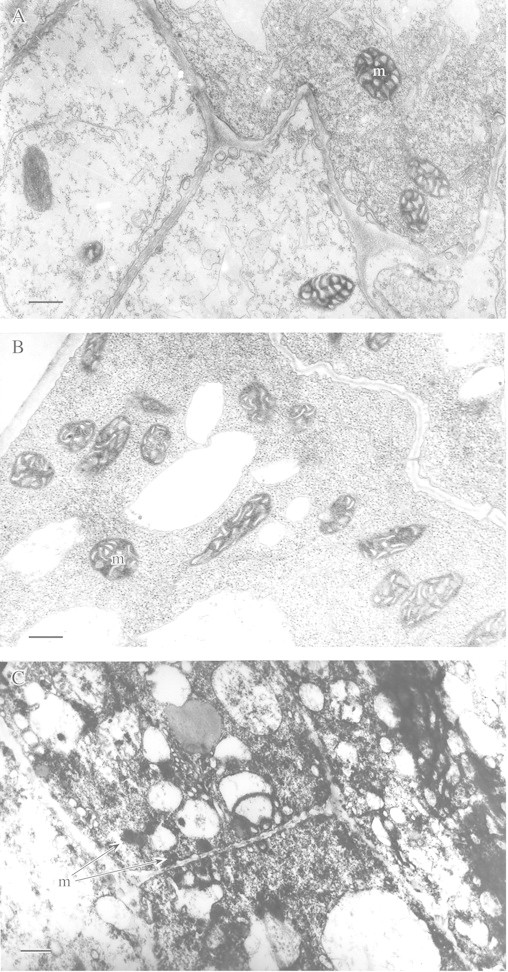
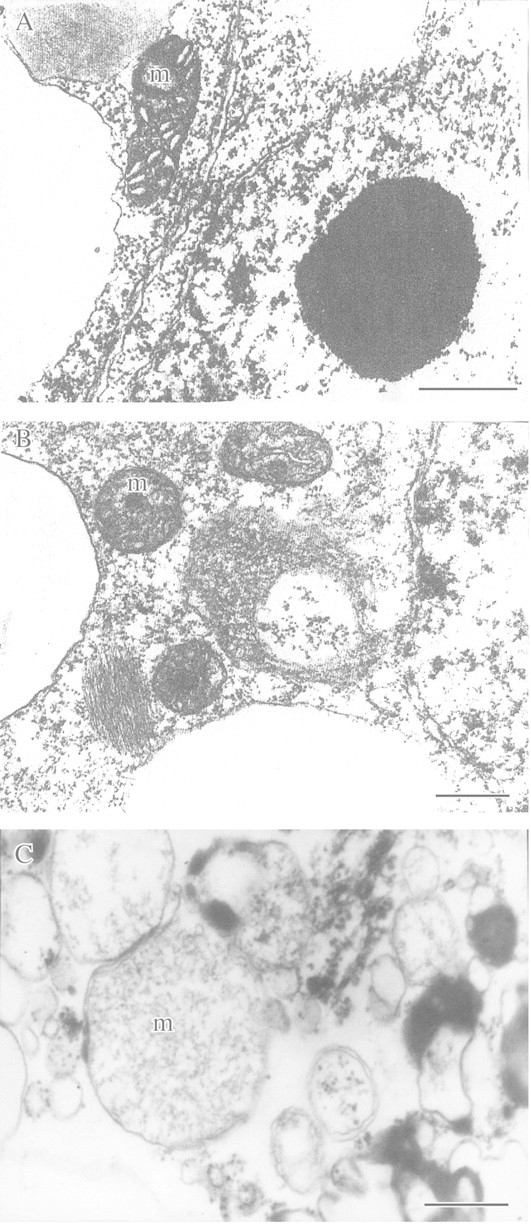
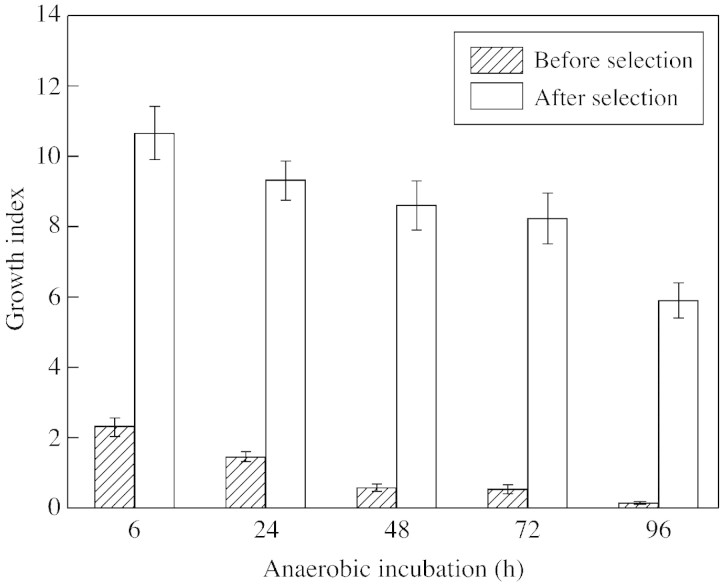

References
-
- AldrichHC, Ferl RJ, Hils MH, Akin DE.1985. Ultrastructural correlates of anaerobic stress in corn roots. Tissue and Cell 17: 341–348. - PubMed
-
- AndreevVYu, Generozova IP, Vartapetian BB.1991. Energy status and mitochondrial ultrastructure of excised pea root at anoxia and postanoxia. Plant Physiology and Biochemistry 29: 171–176.
-
- AndreevaIN, Grineva GM.1970. Ultrastructure of endoplasmic reticulum in the cells of maize root under conditions of hypoxia. Plant Physiology 17: 956–961 (in Russian).
-
- AndrewsCJ.1997. A comparison of glycolytic activity in wheat and two forage grasses in relation to their tolerance to ice encasement. Annals of Botany 79 (Suppl. A): 87–92.
-
- AndrewsCJ, Pomeroy MK.1981. The effect of flooding pretreatment on cold hardiness and survival of winter cereals in ice encasement. Canadian Journal of Plant Science 61: 507–513.
