

Review
doi: 10.1038/nrm1005.
Entry of viruses through the epithelial barrier: pathogenic trickery
Affiliations
- PMID: 12511869
- PMCID: PMC7097689
- DOI: 10.1038/nrm1005
Item in Clipboard
Review
Entry of viruses through the epithelial barrier: pathogenic trickery
Nat Rev Mol Cell Biol.
2003 Jan.
Abstract
Mucosal surfaces--such as the lining of the gut or the reproductive tract--are the main point of entry for viruses into the body. As such, almost all viruses interact with epithelial cells, and make use of the normal epithelial signalling and trafficking pathways of the host cell. In addition to protein receptors, carbohydrate chains of proteoglycans and epithelial-membrane glycosphingolipids have emerged as a new class of receptors for viral attachment to the host cell.
Figures
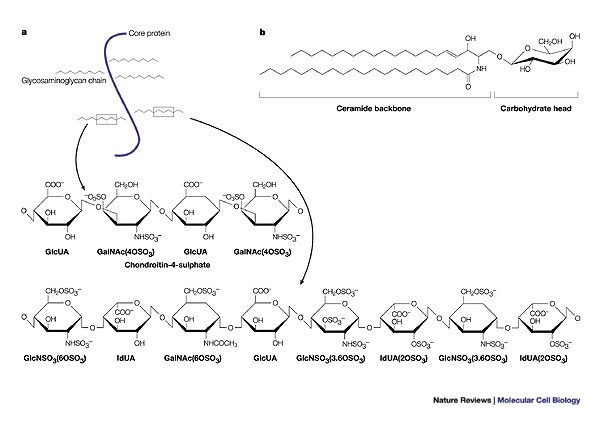
a | Proteoglycans are proteins that are classified by a post-translational attachment of polysaccharide glycosaminoglycan moieties, which are each composed of repeating disaccharide units. The ubiquitously expressed glycosaminoglycan heparan sulphate (shown here) is highly polymorphic, and its sulphated structural motifs are primarily responsible for its protein binding and regulatory properties. b | Glycosphingolipids are anchored in the outer leaflet of the plasma-membrane bilayer by their common hydrophobic backbone, ceramide, which consists of a fatty-acid chain linked to the sphingosine base. The hydrophilic carbohydrate parts of neutral glycosphingolipids and gangliosides protrude into the extracellular space and partially cover the cell surface. GalNAc, (OSO3), N-acetyl galactosamine sulphate; GlcNSO3, glucosamine sulphate; GlcUA, glucuronic acid; IdUA (OSO3), iduronic acid sulphate.

Cell-free human immunodeficiency virus (HIV) or infected cells encounter a mucosal surface (Box 4). Both contain viruses that use either the chemokine receptor CCR5 (R5 virus; found in most acutely infected patients, and therefore thought to be the main vector of infection) or CXCR4 (X4 virus; found later in patients, as the disease progresses) for fusion and/or infection. In the upper small intestine, galactosyl-ceramide (GalCer)+/CCR5+/CXCR4− epithelial cells endocytose cell-free R5 virus at the lumenal surface in a galactosyl ceramide/CCR5-receptor-mediated mechanism, whereas cell-free X4 viruses cannot enter these CXCR4– epithelial cells. Alternatively, R5 (or X4)-infected MONONUCLEAR CELLS bind to the epithelial cell and induce the polarized budding of newly formed R5 (or X4) viruses, which are rapidly endocytosed through GalCer that is present in raft microdomains,. HIV transcytoses across epithelial cells to the serosal surface, where fusion of the transcytotic vesicles releases virus into the LAMINA PROPRIA. Here, R5 viruses infect the CCR5+/CXCR4+/CD4+ lamina propria T lymphocyte, but not the CCR5−/CXCR4−/CD4+ intestinal macrophage. By contrast, the infection of intestinal CXCR4+/CD4+ T cells by X4 viruses that are transcytosed from X4-infected cells is blocked by stromal-cell-derived factor (SDF)-1 that is present in the mucosa. The ability of primary human M cells to translocate R5 or X4 HIV remains to be clarified.

During the first cycle of rotavirus replication in mucosal epithelial cells, the synthesis of rotaviral proteins in the cell cytoplasm leads to an increase in the plasma-membrane permeability to Ca2+, to activation of regulatory mechanisms and to an increase in the concentration of Ca2+ in the endoplasmic reticulum (ER). The increased concentration of cytosolic Ca2+ in infected cells promotes the activation of Ca2+-dependent enzymes, which in turn induces cell lysis and the release of viral proteins and viral progeny. Non-structural protein (NSP)-4 might act as a viral enterotoxin on as-yet-uninfected cells to induce secretory diarrhoea through a | Ca2+-dependent secretion by intestinal cells, b | Ca2+-dependent secretion of peptides and amines to stimulate the enteric nervous system (ENS), and c | further activation of epithelial-cell chloride (Cl−) secretion by the ENS. In parallel, released virus infects downstream absorptive cells. This will lead to a massive cell death and, as a consequence, reduction of the absorptive surface of the intestinal epithelium and an osmotic component of diarrhoea. Various forms of the virus along the rotavirus-maturation pathway are shown: DLP, double-layer particle; IMP, intramembrane particle; Ins(1,4,5)P3, inositol 1,4,5-trisphosphate; PLC, phospholipase C; SERCA, sarcoplasmic/endoplasmic-reticulum Ca2+-ATPase; TLP, triple-layer particle.
References
-
- Gabius H, Andre S, Kaltner H, Siebert H. The sugar code: functional lectinomics. Biochim. Biophys. Acta. 2002;19:2–3. - PubMed
-
- Compans RW. Virus entry and release in polarized epithelial cells. Curr. Top. Microbiol. Immunol. 1995;202:209–219. - PubMed
-
- Bomsel M. Transcytosis of infectious human immunodeficiency virus across a tight human epithelial cell line barrier. Nature Med. 1997;3:42–47. - PubMed
-
- Meng G, et al. Primary intestinal epithelial cells selectively transfer R5 HIV-1 to CCR5+ cells. Nature Med. 2002;8:150–156. - PubMed
-
- Smith GA, Enquist LW. Break ins and break outs: viral interactions with the cytoskeleton of mammalian cells. Annu. Rev. Cell Dev. Biol. 2002;18:135–161. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
