

Catalytic mechanism of thiol peroxidase from Escherichia coli. Sulfenic acid formation and overoxidation of essential CYS61
- PMID: 12514184
- PMCID: PMC3845838
- DOI: 10.1074/jbc.M209888200
Catalytic mechanism of thiol peroxidase from Escherichia coli. Sulfenic acid formation and overoxidation of essential CYS61
Abstract
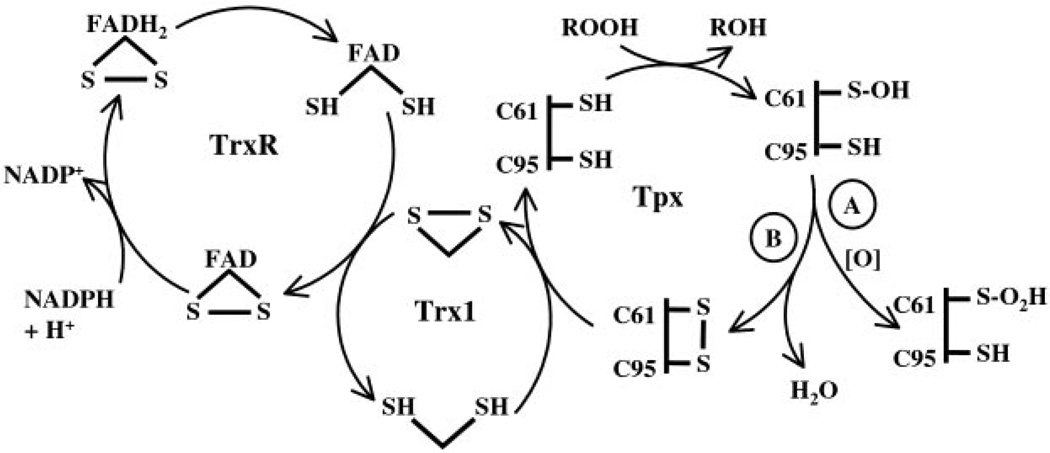
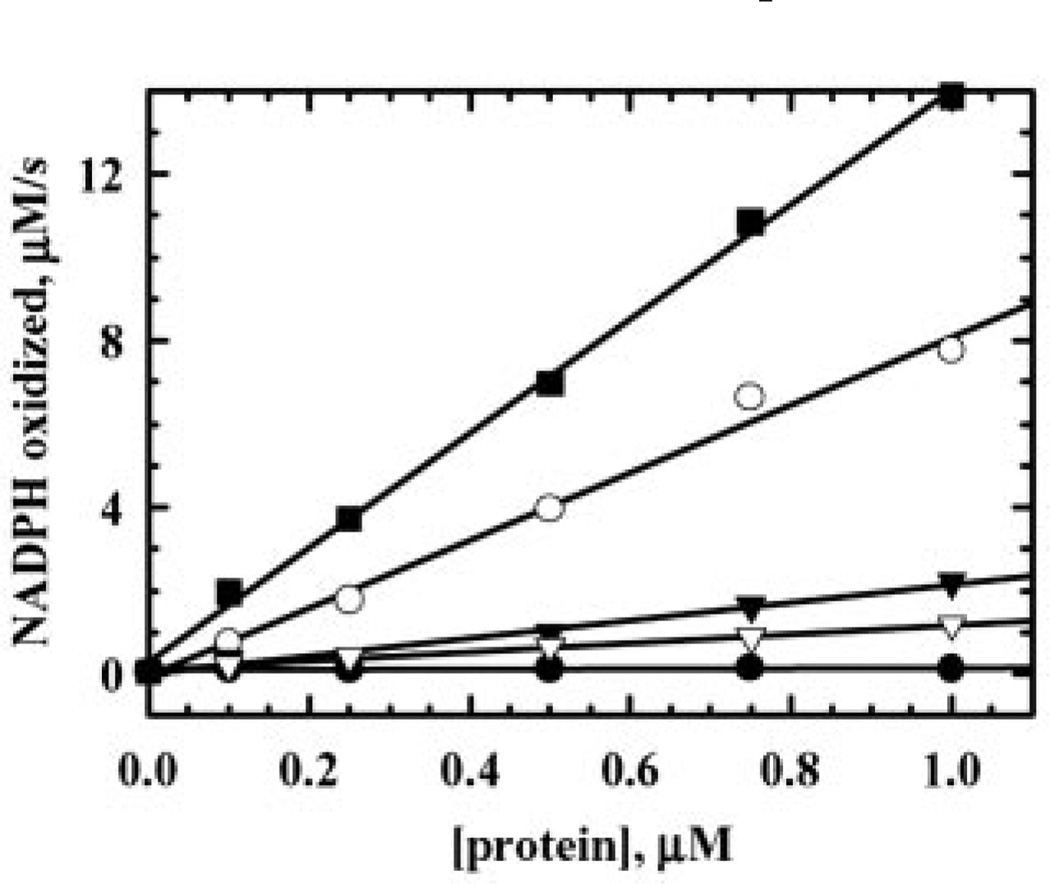
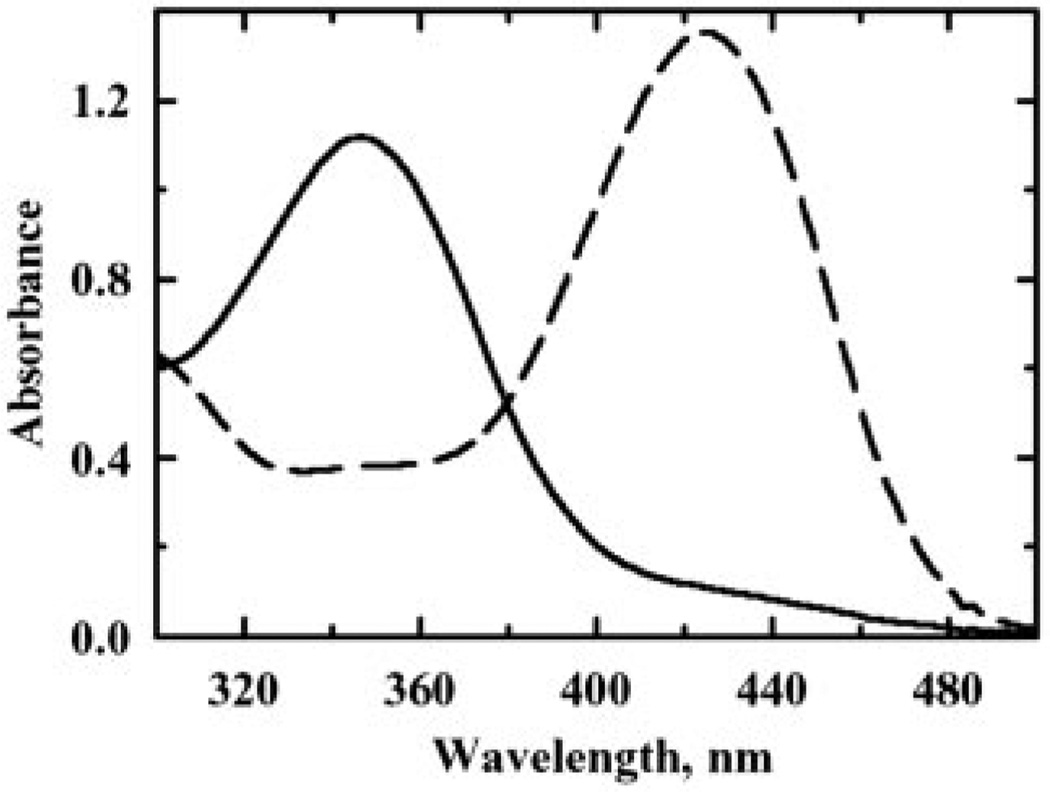
Escherichia coli thiol peroxidase (Tpx, p20, scavengase) is part of an oxidative stress defense system that uses reducing equivalents from thioredoxin (Trx1) and thioredoxin reductase to reduce alkyl hydroperoxides. Tpx contains three Cys residues, Cys(95), Cys(82), and Cys(61), and the latter residue aligns with the N-terminal active site Cys of other peroxidases in the peroxiredoxin family. To identify the catalytically important Cys, we have cloned and purified Tpx and four mutants (C61S, C82S, C95S, and C82S,C95S). In rapid reaction kinetic experiments measuring steady-state turnover, C61S is inactive, C95S retains partial activity, and the C82S mutation only slightly affects reaction rates. Furthermore, a sulfenic acid intermediate at Cys(61) generated by cumene hydroperoxide (CHP) treatment was detected in UV-visible spectra of 4-nitrobenzo-2-oxa-1,3-diazole-labeled C82S,C95S, confirming the identity of Cys(61) as the peroxidatic center. In stopped-flow kinetic studies, Tpx and Trx1 form a Michaelis complex during turnover with a catalytic efficiency of 3.0 x 10(6) m(-1) s(-1), and the low K(m) (9.0 microm) of Tpx for CHP demonstrates substrate specificity toward alkyl hydroperoxides over H(2)O(2) (K(m) > 1.7 mm). Rapid inactivation of Tpx due to Cys(61) overoxidation is observed during turnover with CHP and a lipid hydroperoxide, 15-hydroperoxyeicosatetraenoic acid, but not H(2)O(2). Unlike most other 2-Cys peroxiredoxins, which operate by an intersubunit disulfide mechanism, Tpx contains a redox-active intrasubunit disulfide bond yet is homodimeric in solution.
Figures



References
-
- Storz G, Tartaglia LA, Farr SB, Ames BN. Trends Genet. 1990;6:363–368. - PubMed
-
- Carmel-Harel O, Storz G. Annu. Rev. Microbiol. 2000;54:439–461. - PubMed
-
- Halliwell B, Gutteridge JMC. Free Radicals in Biology and Medicine. 3rd Ed. Oxford: Oxford University Press; 1999. pp. 188–276.
-
- Storz G, Imlay JA. Curr. Opin. Microbiol. 1999;2:188–194. - PubMed
-
- Lynch AS, Lin ECC. In: E. coli and Salmonella: Cellular and Molecular Biology VI. 2nd Ed. Neidhardt FC, editor. Washington, D. C.: American Society for Microbiology Press; 1996. pp. 1526–1538.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
