

Normal female sexual development requires neuregulin-erbB receptor signaling in hypothalamic astrocytes
- PMID: 12514220
- PMCID: PMC6742140
- DOI: 10.1523/JNEUROSCI.23-01-00230.2003
Normal female sexual development requires neuregulin-erbB receptor signaling in hypothalamic astrocytes
Abstract
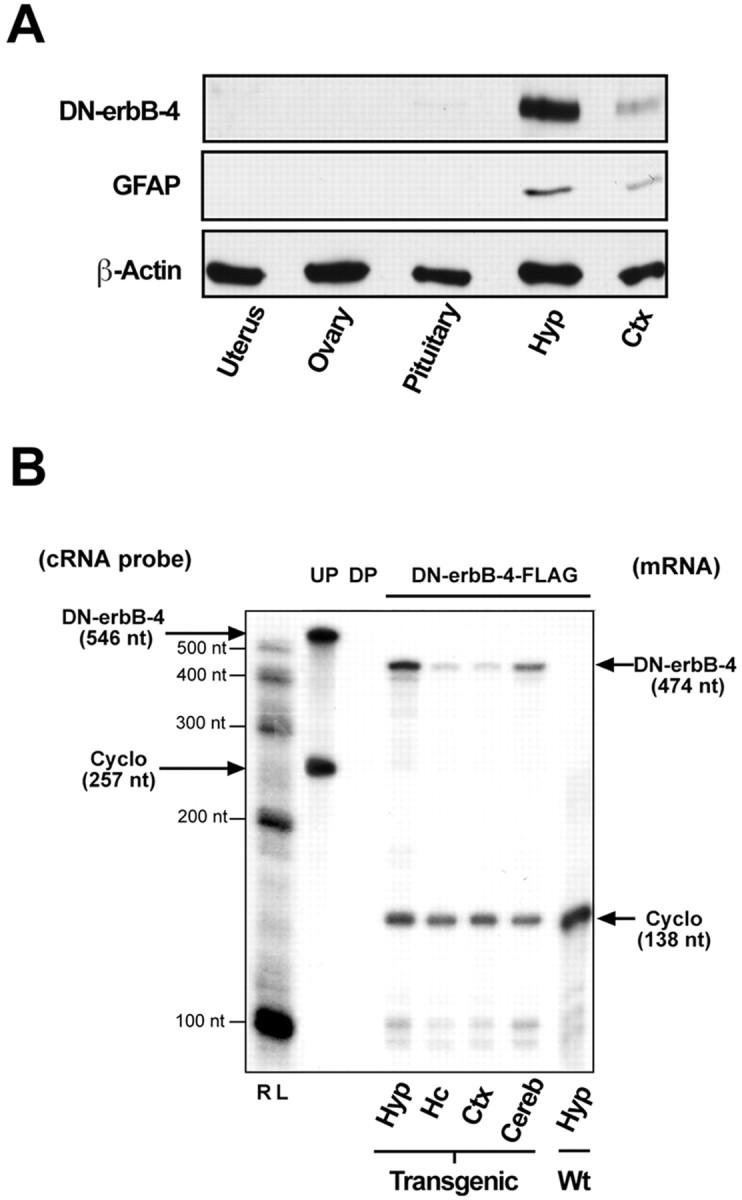
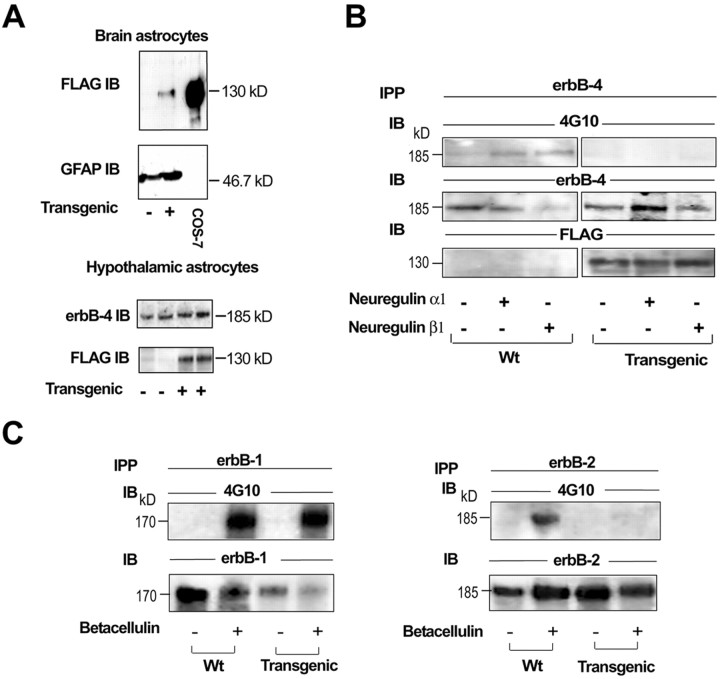
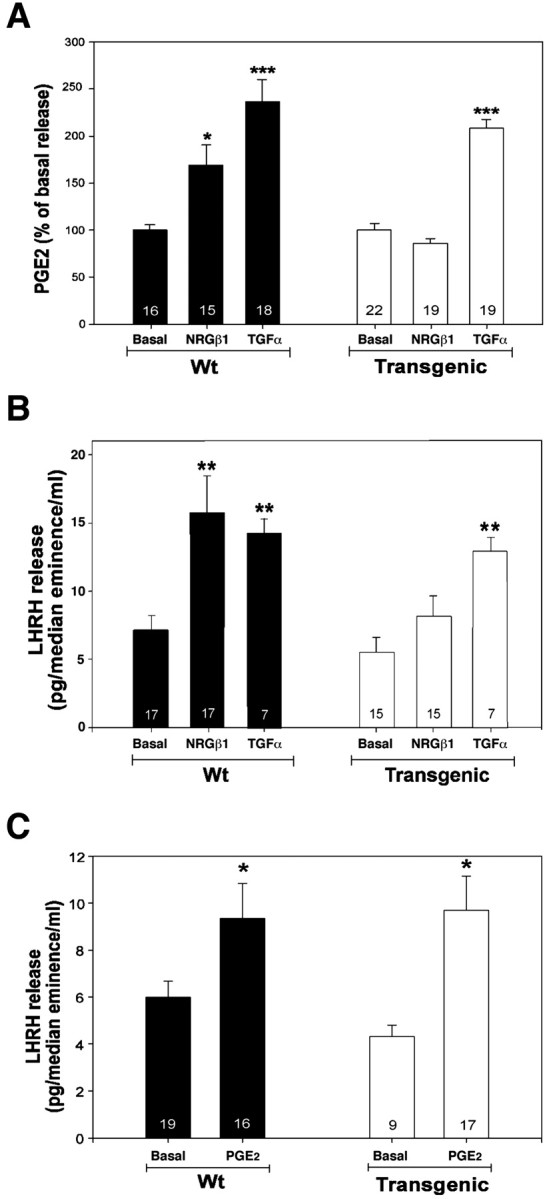
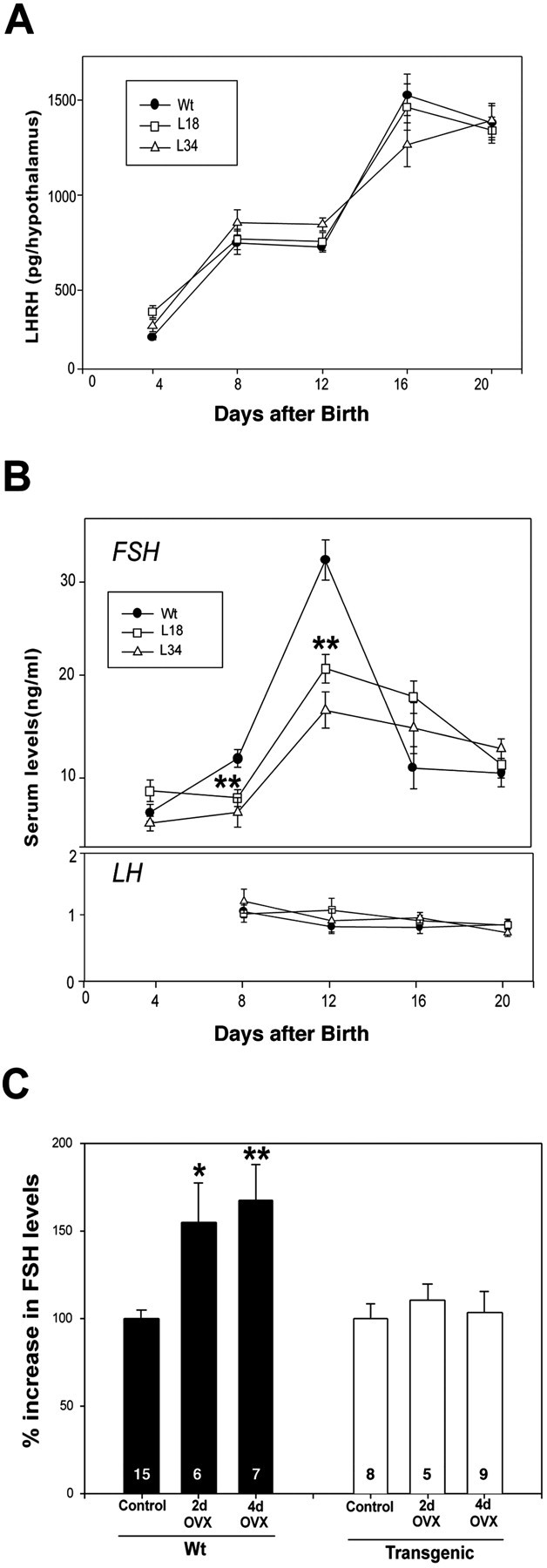
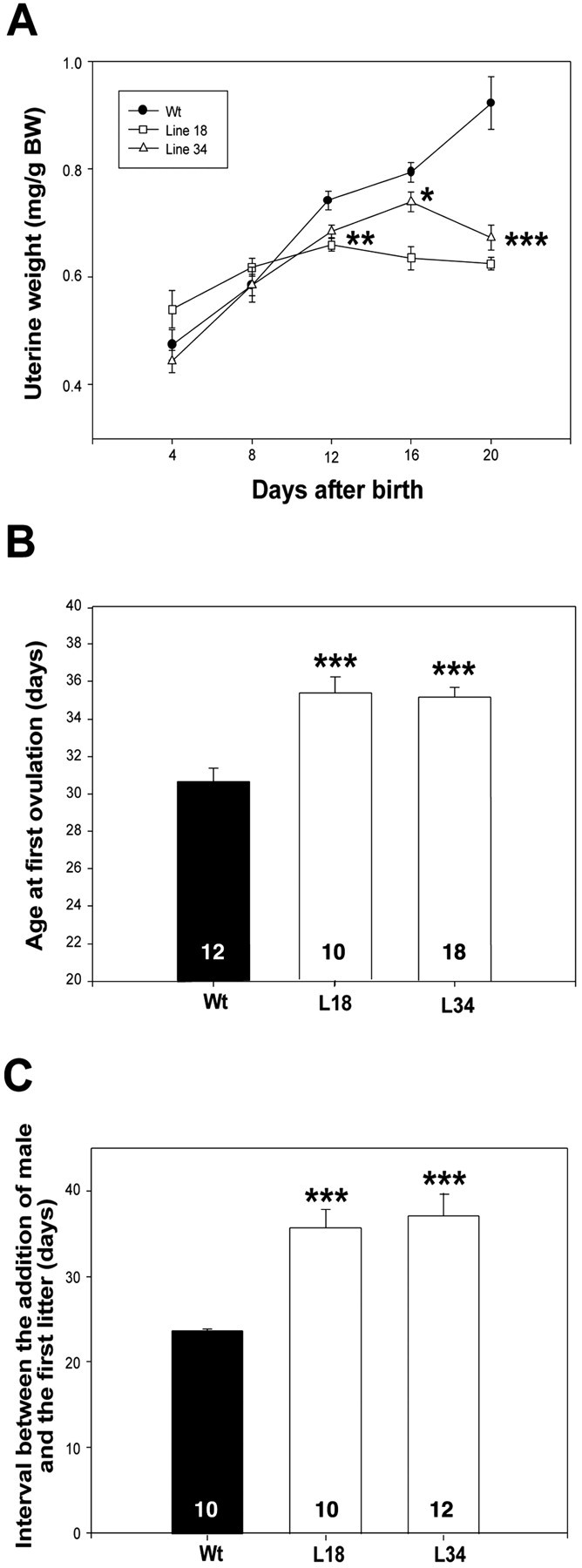
The initiation of mammalian puberty requires the activation of hypothalamic neurons secreting the neuropeptide luteinizing hormone-releasing hormone (LHRH). It is thought that this activation is caused by changes in trans-synaptic input to LHRH neurons. More recently, it has been postulated that the pubertal increase in LHRH secretion in female animals also requires neuron-glia signaling mediated by growth factors of the epidermal growth factor (EGF) family and their astrocytic erbB receptors. Although it appears clear that functional astrocytic erbB1 receptors are necessary for the timely advent of puberty, the physiological contribution that erbB4 receptors may make to this process has not been established. To address this issue, we generated transgenic mice expressing a dominant-negative erbB4 receptor (DN-erbB4) under the control of the GFAP promoter, which targets transgene expression to astrocytes. DN-erbB4 expression is most abundant in hypothalamic astrocytes, where it blocks the ligand-dependent activation of glial erbB4 and erbB2 receptors, without affecting erbB1 (EGF) receptor signaling. Mice carrying the transgene exhibit delayed sexual maturation and a diminished reproductive capacity in early adulthood. These abnormalities are related to a deficiency in pituitary gonadotropin hormone secretion, caused by impaired release of LHRH, the hypothalamic neuropeptide that controls sexual development. In turn, the reduction in LHRH release is caused by the inability of hypothalamic astrocytes to respond to neuregulin (NRG) with production of prostaglandin E(2), which in wild-type animals mediates the stimulatory effect of astroglial erbB receptor activation on neuronal LHRH release. Thus, neuron-astroglia communication via NRG-erbB4/2 receptor signaling appears to be essential for the timely unfolding of the developmental program by which the brain controls mammalian sexual maturation.
Figures
References
-
- Besnard F, Brenner M, Nakatani Y, Chao R, Purohit HJ, Freese E. Multiple interacting sites regulate astrocyte-specific transcription of the human gene for glial fibrillary acidic protein. J Biol Chem. 1991;266:18877–18883. - PubMed
-
- Campbell WB, Ojeda SR. Measurement of prostaglandins by radioimmunoassay. Methods Enzymol. 1987;141:323–350. - PubMed
-
- Chen MS, Bermingham-McDonogh O, Danehy FT, Jr, Nolan C, Scherer SS. Expression of multiple neuregulin transcripts in postnatal rat brains. J Comp Neurol. 1994;349:389–400. - PubMed
-
- Corfas G, Rosen KM, Aratake H, Krauss R, Fischbach GD. Differential expression of ARIA isoforms in the rat brain. Neuron. 1995;14:103–115. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous