

The esterase and PHD domains in CR1-like non-LTR retrotransposons
- PMID: 12519904
- PMCID: PMC7528939
- DOI: 10.1093/molbev/msg011
The esterase and PHD domains in CR1-like non-LTR retrotransposons
Abstract
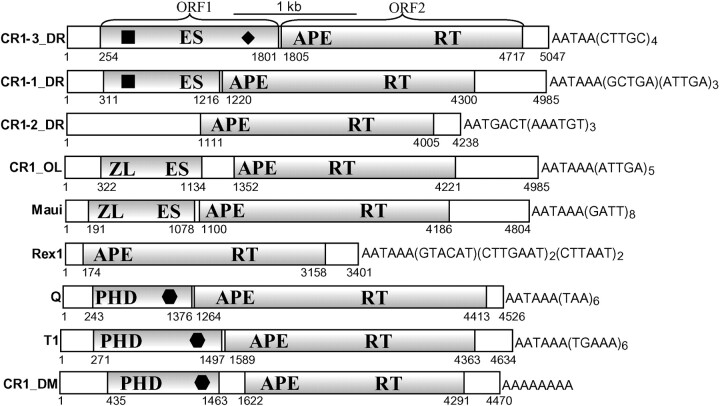
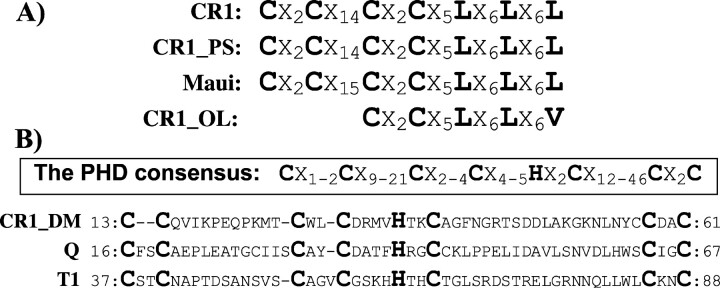
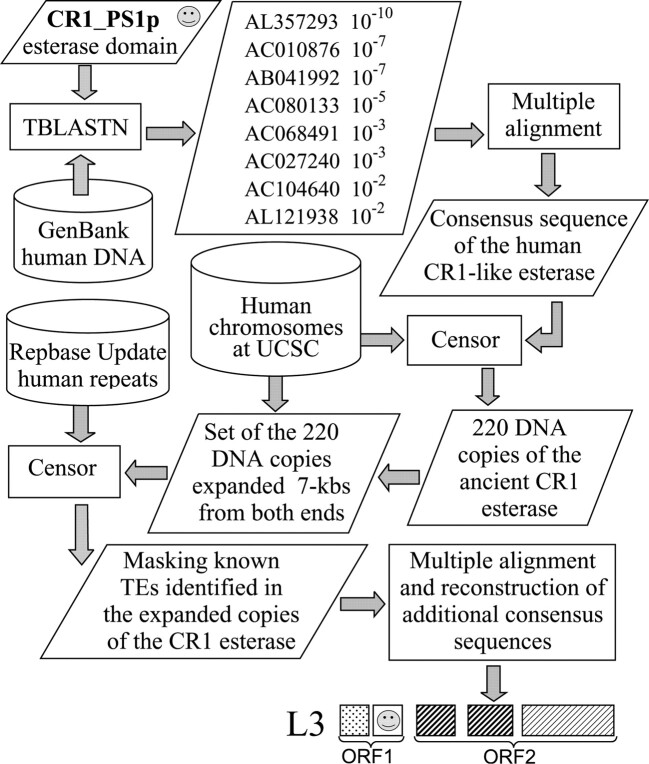
Most active non-LTR (long terminal repeat) retrotransposons carry two open reading frames (ORFs) encoding ORF1p and ORF2p proteins. The ORF2p proteins are relatively well studied and are known to contain endonuclease/reverse transcriptase domains. At the same time, the biological function of ORF1p proteins remains poorly understood, except in that they nonspecifically bind single-stranded mRNA/DNA molecules. CR1-like elements form the most widely distributed clade/superfamily of non-LTR retrotransposons. We found that ORF1p proteins encoded by diverse CR1-like elements contain conserved esterase domain (ES) or plant homeodomain (PHD). This indicates that CR1-like ORF1p proteins are either lipolytic enzymes or are involved in protein-protein interactions related to chromatin remodeling. Sequence conservation of ES suggests that interaction with cellular membranes is an important phase in life circles of CR1-like elements. Presumably such interaction helps in penetrating host cells. As a consequence, the presence of multiple young CR1 families characterized by approximately 10% intrafamily and 40% interfamily identities may be explained by a relatively frequent horizontal transfer of these CR1-like elements. Unexpectedly, ES links together non-LTR retrotransposons and single-stranded RNA viruses like influenza C and coronaviruses, which are known to depend on their own ES.
Figures


References
-
- Aasland, R., T. J. Gibson, and A. F. Stewart. 1995. The PHD finger: implications for chromatin-mediated transcriptional regulation. Trends Biochem. Sci 20:56-59. - PubMed
-
- Bailey, T. L., and W. N. Grundy. 1999. Classifying proteins by family using the product of correlated p-values, pp. 10–14. in P. Istrail, P. Pevzner, and M. Waterman, eds. Proceedings of the Third International Conference on Computational Molecular Biology (RECOMB99). ACM, New York.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
