

doi: 10.1126/science.1079477.
Regulation of blood and lymphatic vascular separation by signaling proteins SLP-76 and Syk
Affiliations
- PMID: 12522250
- PMCID: PMC2982679
- DOI: 10.1126/science.1079477
Item in Clipboard
Regulation of blood and lymphatic vascular separation by signaling proteins SLP-76 and Syk
Science.
.
Abstract
Lymphatic vessels develop from specialized endothelial cells in preexisting blood vessels, but the molecular signals that regulate this separation are unknown. Here we identify a failure to separate emerging lymphatic vessels from blood vessels in mice lacking the hematopoietic signaling protein SLP-76 or Syk. Blood-lymphatic connections lead to embryonic hemorrhage and arteriovenous shunting. Expression of slp-76 could not be detected in endothelial cells, and blood-filled lymphatics also arose in wild-type mice reconstituted with SLP-76-deficient bone marrow. These studies reveal a hematopoietic signaling pathway required for separation of the two major vascular networks in mammals.
Figures
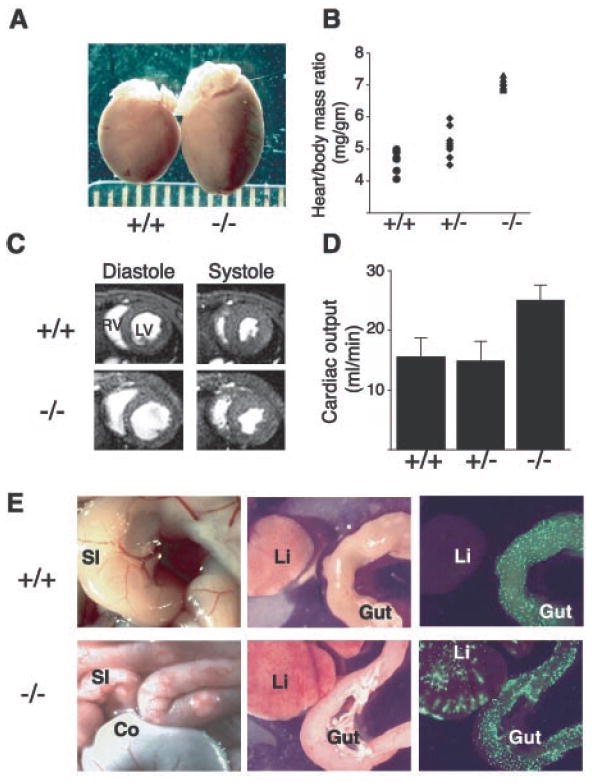
SLP-76–deficient mice exhibit cardiomegaly and elevated cardiac outputs due to A-V shunting of blood. (A) Gross cardiomegaly in a slp-76−/− heart (−/−) compared with that of a wild-type (+/+) littermate at age 2 months. (B) Cardiac hypertrophy in slp-76−/− mice (−/−) is manifest by an elevated heart/body mass ratio compared with wild-type (+/+) and heterozygous (+/−) mice (mean ± standard deviation: +/+ = 4.6 ± 0.4 mg/g; +/− = 5.3 ± 0.5 mg/g; −/− = 7.2 ± 0.4 mg/g; n = 8 animals each; P < 0.0001 for +/+ versus −/−). (C) Cardiac MRI reveals dilated chambers in slp-76−/− hearts with normal ventricular function. Representative cross sections through wild-type (+/+) and slp-76−/− (−/−) hearts at the level of the papillary muscles are shown. Note the larger size of the left (LV) and right (RV) ventricular chambers in the slp-76−/− heart at peak relaxation (diastole) but normal residual volumes at peak contraction (systole). (D) slp-76−/− mice have markedly elevated resting cardiac outputs. Cardiac outputs of slp-76+/+ (+/+), slp-76+/− (+/−), and slp-76−/− (−/−) mice were measured by MRI as described (mean ± standard deviation: +/+ = 15.6 ± 3.1 ml/min; +/− = 15.0 ± 3.2 ml/min; −/− = 25.1 ± 2.5 ml/min; n = 5 animals each; P < 0.001 for +/+ versus −/−). (E) A-V shunting in slp-76−/− mice. slp-76−/− mice (−/−) have dilated, tortuous blood vessels in the small intestine (SI) but not in the colon (Co) (left). Fluorescent microspheres (15 μm) injected into the aorta pass through the intestinal circulation to lodge in the liver (Li) of slp-76−/− (−/−) but not wild-type (+/−) mice (right).
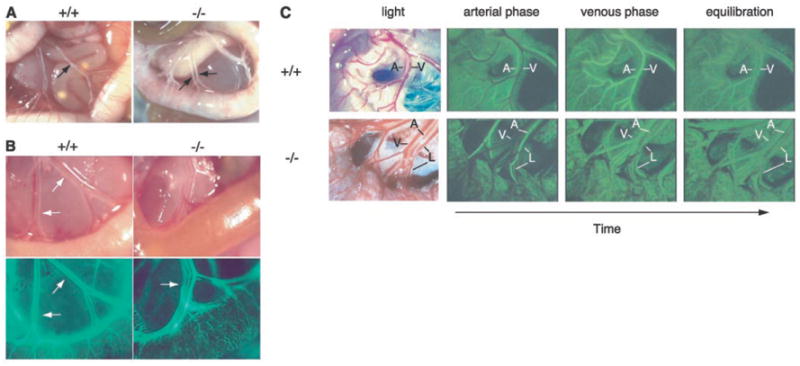
Vascular malformations in the intestine of neonatal slp-76−/− mice mediate mixing of lymphatic and blood circulations. (A) Chyle-filled mesenteric blood vessels (arrows) are present in a slp-76−/− mouse (−/−) in association with a gut wall that lacks normal external blood vessels. (B) FITC-dextran injection demonstrates communication between the blood and lymphatic circulations in neonatal slp-76−/− mice. Fluorescent imaging of mesenteric vessels after left ventricular injection of FITC-dextran identifies the lymphatics (arrow) in slp-76−/− but not wild-type +/+ neonates. (C) First-pass FITC-dextran angiography of adult slp-76−/− mice demonstrates vascular continuity between mesenteric arteries, veins, and lymphatics. Real-time images of the mesenteric vasculature of live wild-type (+/+) and slp-76−/− (−/−) mice were obtained after intravenous injection of FITC-dextran. The artery (A) fills first in both wild-type and slp-76−/− mice, followed by the vein (V) in wild-type mice and by simultaneous filling of both the vein and lymphatic (L) in slp-76−/− mice. Images were obtained at peak intensity for the artery (arterial phase), peak intensity for the vein (venous phase), and after complete mixing of the FITC-dextran in the circulation (equilibration).
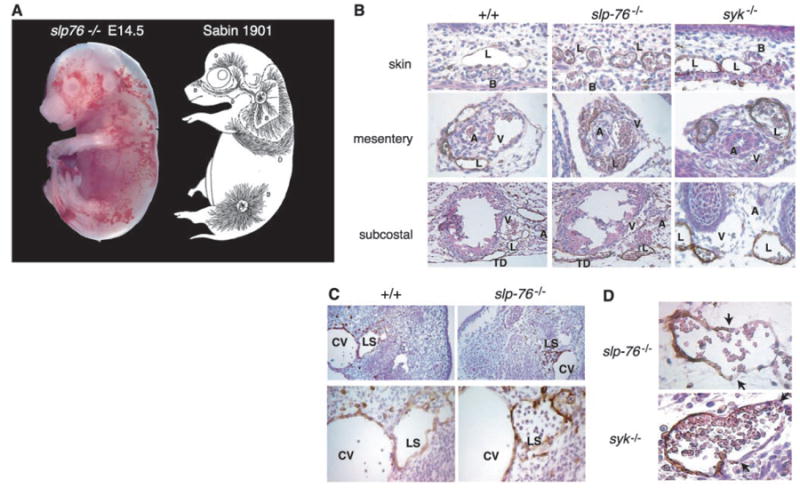
The developing lymphatic circulation in mice lacking SLP-76 and Syk communicates with the blood circulation. (A) The pattern of superficial vascularity and hemorrhage in an E14.5 slp-76−/− embryo (left) is strikingly similar to that of the developing superficial lymphatic vessels described by Sabin (3) (right, reproduced with permission of Wiley Periodicals, Inc.). (B) Developing lymphatic vessels in slp-76−/− and syk−/− embryos are filled with blood. LYVE staining in the skin, mesentery, and subcostal region reveals blood-filled lymphatics in E15.5 slp-76−/− and syk−/− but not wild-type (+/+) embryos. B, blood vessel; L, lymphatic vessel; A, artery; V, vein; TD, thoracic duct. Note that the number and distribution of lymphatics in slp-76−/− and syk−/− embryos are similar to those in wild-type embryos. (C) Blood-filled lymph sacs precede hemorrhage in slp-76−/− embryos. LYVE staining of transverse sections of E11.5 wild-type and slp-76−/− embryos is shown at low power (top) and high power (bottom). Blood is present in slp-76−/− lymph sacs before development of true lymphatic vessels and in the absence of hemorrhage. CV, cardinal vein; LS, lymph sac. (D) Identification of blood-lymphatic vascular chimeras in slp-76−/− and syk−/− embryos. LYVE staining of slp-76−/− and syk−/− embryos reveals chimeric vessels composed of lymphatic endothelial cells (LYVE-positive) fused to blood endothelial cells (LYVE-negative). Blood-lymphatic endothelial junctions are demarcated by arrows.
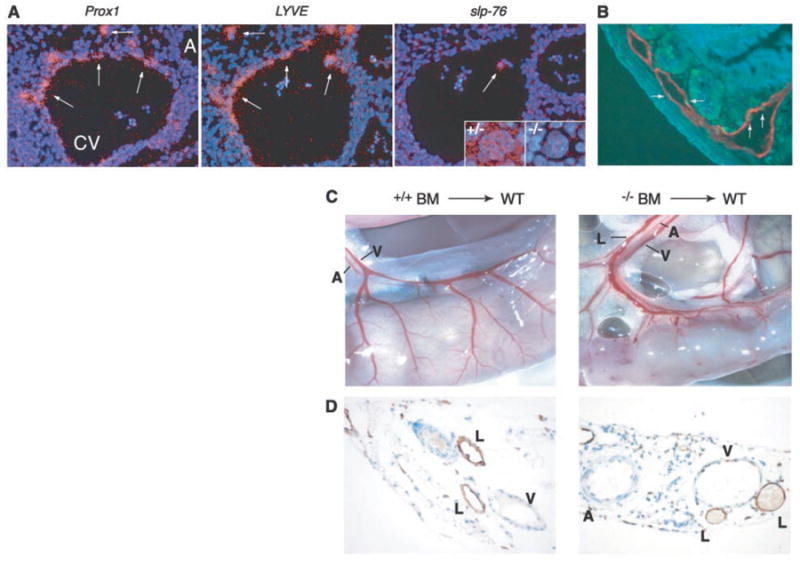
Separation of lymphatic vessels from blood vessels requires SLP-76 signaling in hematopoietic cells. (A) slp-76 is not expressed in lymphatic endothelial progenitor cells. In situ hybridization of wild-type E11.5 embryos with probes for Prox1 and LYVE identifies lymphatic endothelial progenitors (arrows) as they arise from the cardinal vein (left and middle, CV, cardinal vein; A, aorta). slp-76 mRNA cannot be detected in Prox1-positive/LYVE-positive cells but is detected in a single circulating cell within the lumen of the cardinal vein (white arrow) and in megakaryocytes from slp-76+/− (+/−) but not SLP-76–deficient (−/−) fetal liver. (B) Radiation induces lymphatic endothelial apoptosis. Wild-type mice were sacrificed 4 hours after exposure to 1200 rad of total body irradiation and the intestine was stained for both TUNEL (terminal deoxynucleotidyl transferase–mediated dUTP nick-end labeling) (green) and LYVE (red) to identify apoptotic lymphatic endothelial cells (arrows). (C) Transplantation of slp-76−/− bone marrow into lethally irradiated wild-type mice confers a vascular phenotype identical to that of slp-76−/− mice. After transplantation with slp-76−/− but not wild-type bone marrow (BM), the intestine of a wild-type mouse loses normal blood vessel architecture and blood is seen filling three mesenteric vessels rather than two. A, artery, V, vein, L, lymphatic. (D) Mixing of blood and lymphatic circulations after transplantation of slp-76−/− bone marrow into lethally irradiated wild-type mice. LYVE staining of the mesentery of the animals shown in (C) reveals blood-filled lymphatics in the animal reconstituted with SLP-76–deficient but not wild-type marrow.
Comment in
-
Development. Lymphatics make the break.Science. 2003 Jan 10;299(5604):209-10. doi: 10.1126/science.1081345. Science. 2003. PMID: 12522236 No abstract available.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
