

The yeast counterparts of human 'MELAS' mutations cause mitochondrial dysfunction that can be rescued by overexpression of the mitochondrial translation factor EF-Tu
- PMID: 12524521
- PMCID: PMC1315813
- DOI: 10.1038/sj.embor.embor713
The yeast counterparts of human 'MELAS' mutations cause mitochondrial dysfunction that can be rescued by overexpression of the mitochondrial translation factor EF-Tu
Abstract
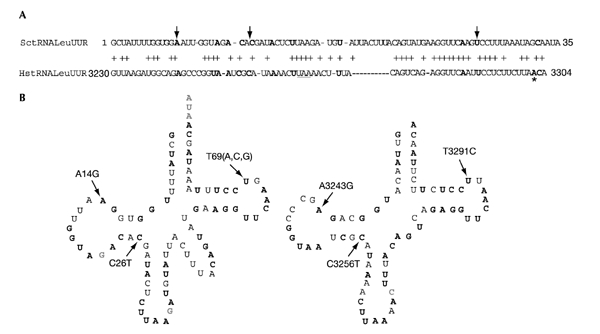
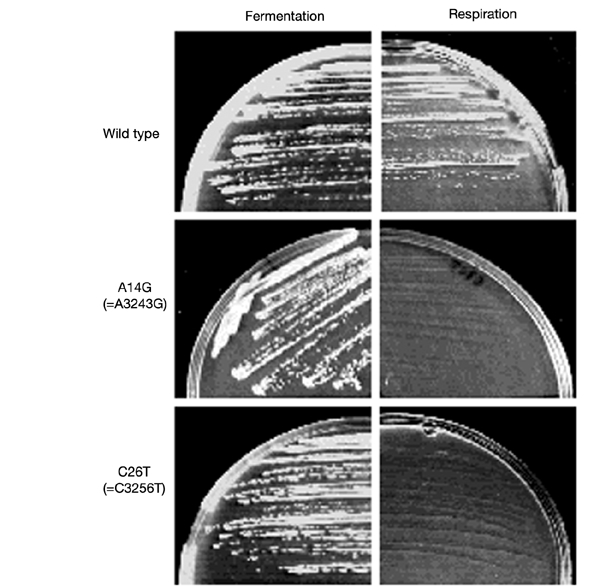
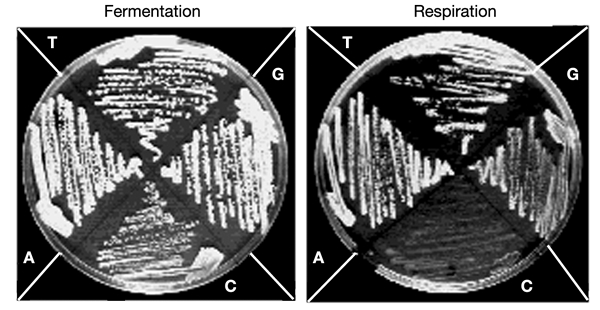
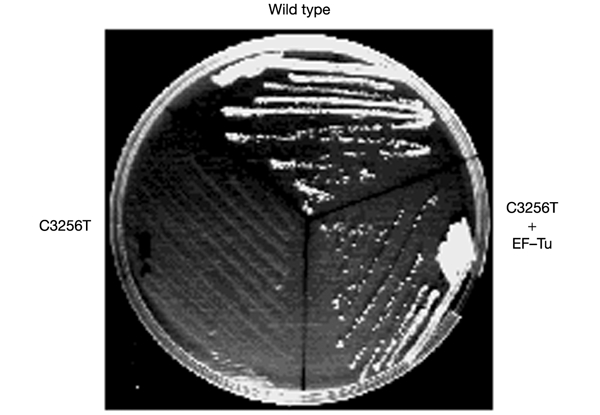
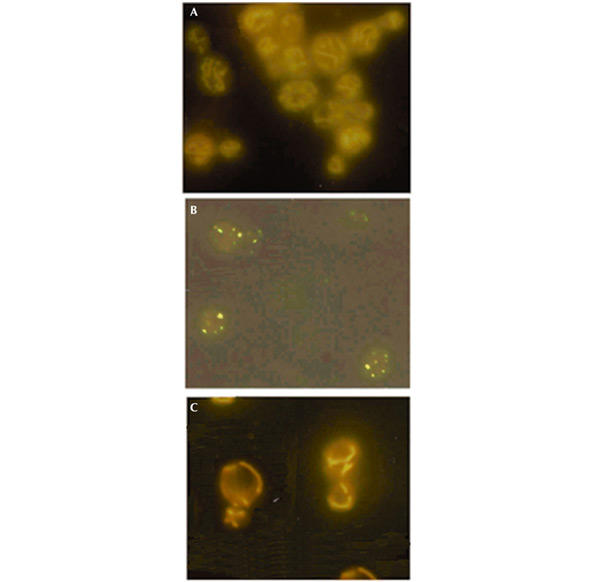
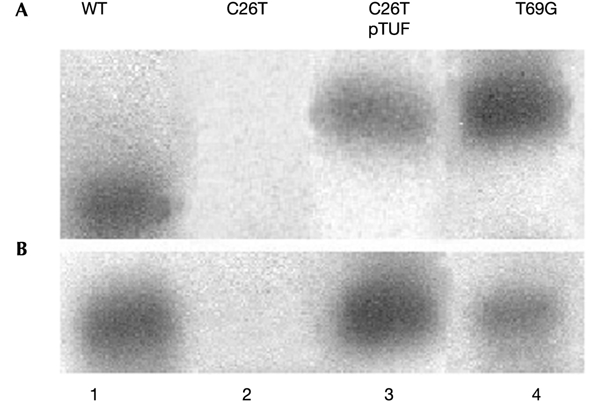
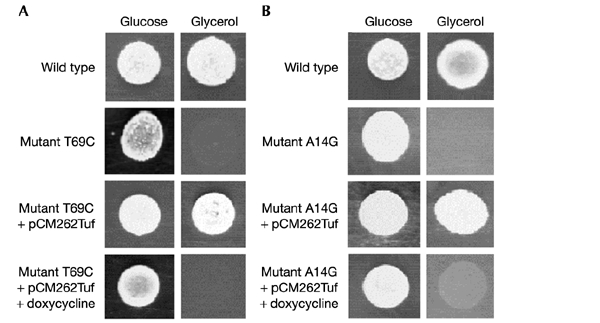
We have taken advantage of the similarity between human and yeast (Saccharomyces cerevisiae) mitochondrial tRNA(Leu)(UUR), and of the possibility of transforming yeast mitochondria, to construct yeast mitochondrial mutations in the gene encoding tRNA(Leu)(UUR) equivalent to the human A3243G, C3256T and T3291C mutations that have been found in patients with the neurodegenerative disease MELAS (for mitochondrial 'myopathy, encephalopathy, lactic acidosis and stroke-like episodes'). The resulting yeast cells (bearing the equivalent mutations A14G, C26T and T69C) were defective for growth on respiratory substrates, exhibited an abnormal mitochondrial morphology, and accumulated mitochondrial DNA deletions at a very high rate, a trait characteristic of severe mitochondrial defects in protein synthesis. This effect was specific at least in the pathogenic mutation T69C, because when we introduced A or G instead of C, the respiratory defect was absent or very mild. All defective phenotypes returned to normal when the mutant cells were transformed by multicopy plasmids carrying the gene encoding the mitochondrial elongation factor EF-Tu. The ability to create and analyse such mutated strains and to select correcting genes should make yeast a good model for the study of tRNAs and their interacting partners and a practical tool for the study of pathological mutations and of tRNA sequence polymorphisms.
Figures
References
-
- Bonnefoy N. & Fox T.D. (2001) Genetic transformation of S. cerevisiae mitochondria. Methods Cell Biol., 65, 381–396. - PubMed
-
- Costanzo M.C. & Fox T.D. (1990) Control of mitochondrial gene expression in Saccharomyces cerevisiae. Annu. Rev. Genet., 24, 91–113. - PubMed
-
- Fox T.D., Folley L.S. & Mulero J.J. (1991) Analysis and manipulation of yeast mitochondrial genes. Methods Enzymol., 194, 149–165. - PubMed
-
- Francisci S., Bohn C., Frontali L. & Bolotin-Fukuhara M. (1998) Ts mutations in mitochondrial tRNA genes: characterization and effects of two point mutations in the mitochondrial gene for tRNAphe in Saccharomyces cerevisiae. Curr. Genet., 33, 110–116. - PubMed
-
- Gari E., Piedrafita L., Aldea M. & Herrero E. (1997) A set of vectors with a tetracycline-regulatable promoter system for modulated gene expression in Saccharomyces cerevisiae. Yeast, 13, 837–848. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
