

Review
doi: 10.1128/jvi.77.3.1659-1665.2003.
Latency in human immunodeficiency virus type 1 infection: no easy answers
Affiliations
- PMID: 12525599
- PMCID: PMC140995
- DOI: 10.1128/jvi.77.3.1659-1665.2003
Item in Clipboard
Review
Latency in human immunodeficiency virus type 1 infection: no easy answers
J Virol.
2003 Feb.
No abstract available
Figures
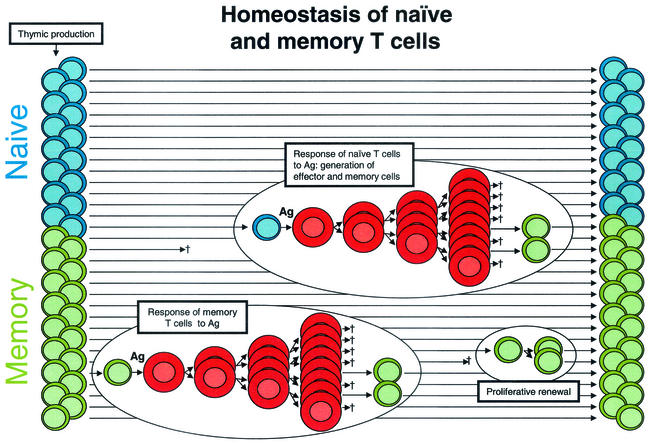
Normal T-cell homeostasis. Most of the CD4+ T cells in the body are small resting cells that circulate throughout the lymphoid tissues, poised to respond to a specific Ag. Approximately half are naïve cells (blue) that have not encountered an Ag since emerging from the thymus. The remainder are memory cells (green) that have previously responded to Ag. Following their encounter with Ag, resting cells undergo blast transformation and begin to proliferate. These lymphoblasts (red) undergo several rounds of cell division, giving rising to effector cells. Most effector cells eventually die, but a fraction revert to a resting memory state. The memory pool is maintained by the long life span of the cells and a gradual process of proliferative renewal.
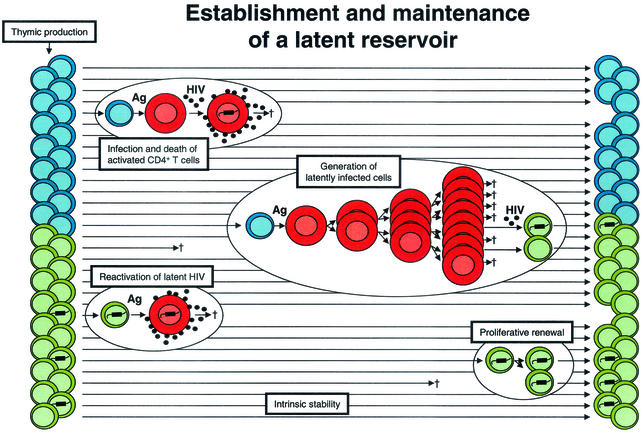
Establishment of a latent reservoir in resting memory CD4+ T cells. CD4+ lymphoblasts (red cells) are highly susceptible to productive infection and usually die within a few days after infection. Latently infected cells with integrated HIV-1 DNA may be generated when lymphoblasts that are in the process of reverting to a resting state become infected. When latently infected cells subsequently encounter the relevant Ag, they become permissive for virus gene expression and virus production. Latently infected cells may be maintained by intrinsic stability, as well as by the process of proliferative renewal if they do not become susceptible to HIV-1-induced cytopathic effects or host cytolytic mechanisms during this process.
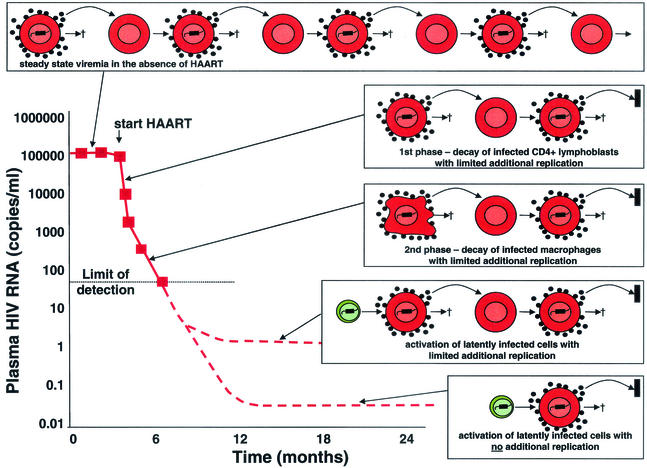
Sources of virus in plasma during HAART. In the absence of treatment, there is an equilibrium between the production and clearance of infected cells. With HAART, there is a rapid decay of plasma virus (t1/2 of hours) and of the infected CD4+ T lymphoblasts that produce most of the plasma virus (t1/2 = 1 day) (23, 55). The second phase of decay results from the slower turnover (t1/2 = 2 weeks) of another population of infected cells, perhaps macrophages (38). The second phase brings the level of viremia down to a new steady state somewhat below the limit of detection of current assays for plasma HIV-1 RNA (50 copies/ml). It was initially assumed that HAART immediately stopped all new infection of susceptible cells (23, 55). More recently, it has been argued that HAART may not completely arrest viral replication but may simply reduce the number of newly infected cells generated from each productively infected cell to substantially below one (18). In patients who are responding well to HAART, there is release of virus from stable reservoirs and some degree of additional replication, giving rise to a new steady-state level of viremia that is <50 copies/ml. However, the additional replication is sufficiently limited that new drug resistance mutations rarely arise in such patients (22). If HAART stopped all new infection of susceptible cells, then virus levels in plasma would fall to a basal level, the magnitude of which would depend on the size and average rate of activation of the latent compartment (with possible contributions from other stable reservoirs).
References
-
- Adams, M., L. Sharmeen, J. Kimpton, J. M. Romeo, J. V. Garcia, B. M. Peterlin, M. Groudine, and M. Emerman. 1994. Cellular latency in human immunodeficiency virus-infected individuals with high CD4 levels can be detected by the presence of promoter-proximal transcripts. Proc. Natl. Acad. Sci. USA 91:3862-3866. - PMC - PubMed
-
- Brooks, D. G., S. G. Kitchen, C. M. Kitchen, D. D. Scripture-Adams, and J. A. Zack. 2001. Generation of HIV latency during thymopoiesis. Nat. Med. 7:459-464. - PubMed
-
- Cavert, W., D. W. Notermans, K. Staskus, S. W. Wietgrefe, M. Zupancic, K. Gebhard, K. Henry, S.-C. Zhang, R. Mills, H. McDade, J. Goudsmit, S. A. Danner, and A. T. Haase. 1997. Kinetics of response in lymphoid tissue to antiretroviral therapy of HIV-1 infection. Science 276:960-964. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
