

doi: 10.1128/jvi.77.3.2287-2294.2003.
Duck hepatitis B virus virion secretion requires a double-stranded DNA genome
Affiliations
- PMID: 12525667
- PMCID: PMC140969
- DOI: 10.1128/jvi.77.3.2287-2294.2003
Item in Clipboard
Duck hepatitis B virus virion secretion requires a double-stranded DNA genome
J Virol.
2003 Feb.
Abstract
Hepatitis B virus assembly begins with the packaging of an RNA pregenome into intracellular nucleocapsids, with subsequent reverse transcription within these nucleocapsids converting the RNA into a characteristic, partially double-stranded DNA, which, alone, is found in enveloped extracellular virions as the viral genome. Using a synchronized replication system for the duck hepatitis B virus, together with a stringent two-step assay for virion secretion, we demonstrate that this selective genome secretion results from an intrinsic secretion competence gained only by the nucleocapsids at the late stage of reverse transcription.
Figures
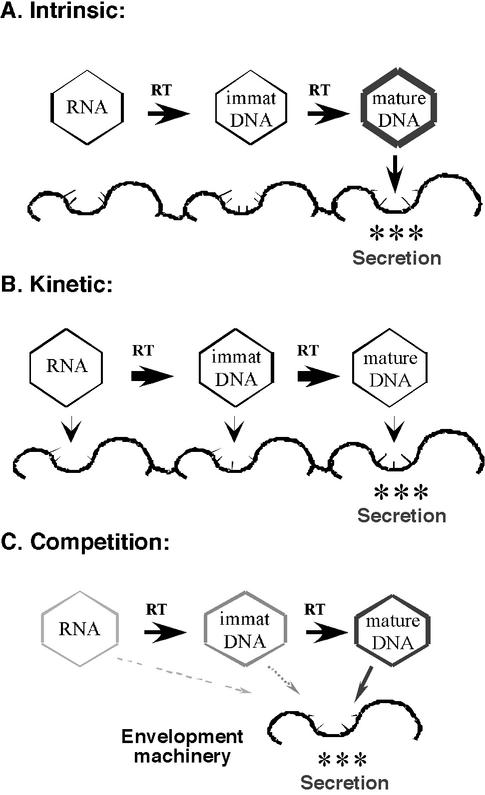
Models of nucleocapsid maturation. Three alternative models explaining the phenomenon of nucleocapsid maturation in hepadnaviruses, i.e., the apparent selective envelopment and secretion of only mature DNA-containing nucleocapsids. (A) The intrinsic model. An intrinsic property (a “maturation signal”) emerges on mature nucleocapsids (represented by the thick capsid shell) that endows them, alone, with secretion competence, i.e., the ability to interact with the secretion machinery (the thick wavy line with thin studs). Immature (immat) nucleocapsid species, containing pgRNA or immature DNA intermediates of reverse transcription (RT), are not secreted because they lack this intrinsic property. (B) The kinetic model. Each species of nucleocapsid has equal intrinsic secretion competence. However, only mature nucleocapsids are secreted because the kinetics of reverse transcription (shown as thick arrows) is faster than the rate of envelopment and secretion (shown as thin arrows), and, therefore, reverse transcription outcompetes the secretion process for the immature nucleocapsids. In other words, all species of nucleocapsids have had the opportunity to complete reverse transcription by the time that they exit the “secretion pipeline.” (C) The competition model. In the context of competition over a limiting secretion machinery, any increased secretion competence and/or increased abundance (represented by the thickening and darkening of the capsid shell) of mature nucleocapsids over the immature species is amplified. Even if the mature nucleocapsids are only slightly more secretion competent and/or abundant, they may outcompete immature nucleocapsids for this limiting secretion machinery.
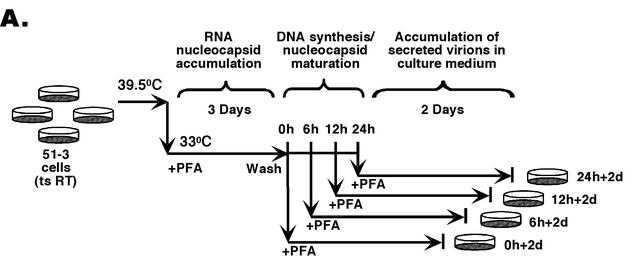
(A) The synchronized replication system. The synchronized DHBV replication system utilizes an LMH-derived cell line, 51-3, that contains a stably integrated DHBV genome harboring a temperature-sensitive (ts) RT mutant, in conjunction with the reversible RT inhibitor, PFA, to synchronize viral replication (4, 21). This system was used to assess the intrinsic secretion competence of immature nucleocapsids, in the absence of ongoing reverse transcription and potential competition from mature nucleocapsids. Cells were expanded and grown to confluence at the nonpermissive temperature (39.5°C) for viral replication and were then shifted to the permissive temperature (33°C) while PFA (final concentration of 1 mM) was added and were maintained at this temperature in the presence of PFA for a period of 3 days to allow for the accumulation of large quantities of pgRNA-containing nucleocapsids. Cells were then washed free of PFA, and DNA synthesis within the nucleocapsids was allowed to proceed in a synchronized fashion for 0, 6, 12, and 24 h. DNA synthesis was then frozen in its progression by the readdition of PFA, and the cells were maintained for an additional 2 days (d) in culture at 33°C in the presence of PFA to allow for the accumulation of secreted virions in the culture supernatants. Culture supernatants were collected both immediately before and after the second stage of PFA treatment to assay for the presence of enveloped virions. (B) Southern blot analysis of intracellular nucleocapsid DNA from the synchronized replication system. Viral DNA was extracted from nucleocapsids that had synthesized viral DNA for 0, 6, 12, and 24 h, respectively, either before (lanes 1 to 4; 0, 6, 12, and 24 h, respectively) or after (lanes 5 to 8; 0 h + 2 days [d], 6 h + 2 days, 12 h + 2 days, and 24 h + 2 days, respectively) the second PFA treatment period (the virion accumulation period) and was detected by Southern blot analysis. RC DNA (RC) is the major form of the mature, ds DNA; ss DNA (SS) is the immature reverse transcription intermediate. These experiments were repeated six times with similar results obtained.
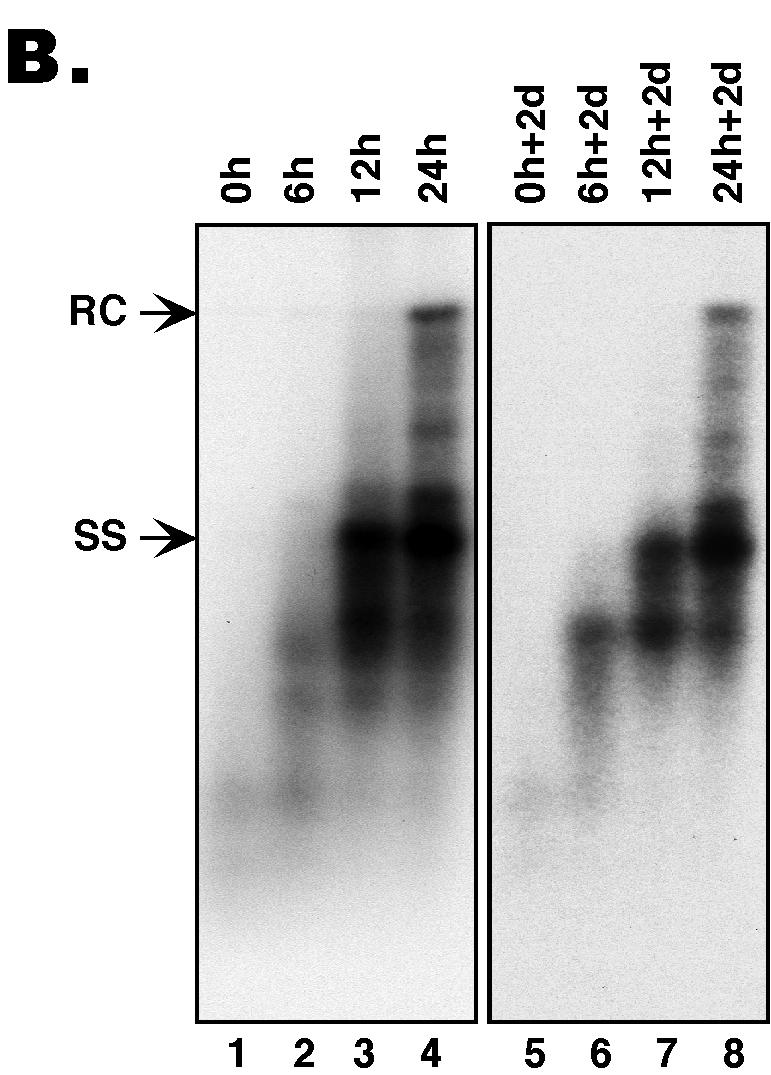
(A) The synchronized replication system. The synchronized DHBV replication system utilizes an LMH-derived cell line, 51-3, that contains a stably integrated DHBV genome harboring a temperature-sensitive (ts) RT mutant, in conjunction with the reversible RT inhibitor, PFA, to synchronize viral replication (4, 21). This system was used to assess the intrinsic secretion competence of immature nucleocapsids, in the absence of ongoing reverse transcription and potential competition from mature nucleocapsids. Cells were expanded and grown to confluence at the nonpermissive temperature (39.5°C) for viral replication and were then shifted to the permissive temperature (33°C) while PFA (final concentration of 1 mM) was added and were maintained at this temperature in the presence of PFA for a period of 3 days to allow for the accumulation of large quantities of pgRNA-containing nucleocapsids. Cells were then washed free of PFA, and DNA synthesis within the nucleocapsids was allowed to proceed in a synchronized fashion for 0, 6, 12, and 24 h. DNA synthesis was then frozen in its progression by the readdition of PFA, and the cells were maintained for an additional 2 days (d) in culture at 33°C in the presence of PFA to allow for the accumulation of secreted virions in the culture supernatants. Culture supernatants were collected both immediately before and after the second stage of PFA treatment to assay for the presence of enveloped virions. (B) Southern blot analysis of intracellular nucleocapsid DNA from the synchronized replication system. Viral DNA was extracted from nucleocapsids that had synthesized viral DNA for 0, 6, 12, and 24 h, respectively, either before (lanes 1 to 4; 0, 6, 12, and 24 h, respectively) or after (lanes 5 to 8; 0 h + 2 days [d], 6 h + 2 days, 12 h + 2 days, and 24 h + 2 days, respectively) the second PFA treatment period (the virion accumulation period) and was detected by Southern blot analysis. RC DNA (RC) is the major form of the mature, ds DNA; ss DNA (SS) is the immature reverse transcription intermediate. These experiments were repeated six times with similar results obtained.
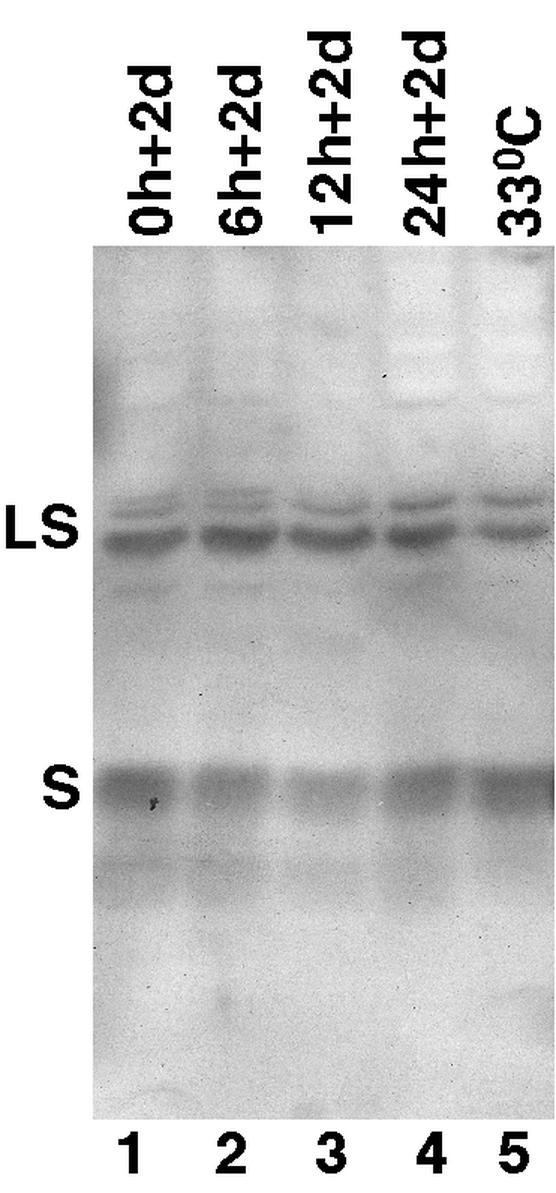
Western blot analysis of viral surface proteins. Cell culture medium was collected from the 51-3 cells at the various time points (Fig. 2) of the synchronized system (lanes 1 to 4) as well as from a steady-state culture maintained at 33°C for 7 days in the absence of PFA (lane 5). The DHBV surface proteins present in the medium samples were then detected by sodium dodecyl sulfate-polyacrylamide gel electrophoresis and Western blot analysis by using a monoclonal antibody specific for the S domain of the DHBV surface proteins (16). The large (LS) and small (S) surface proteins were indicated.
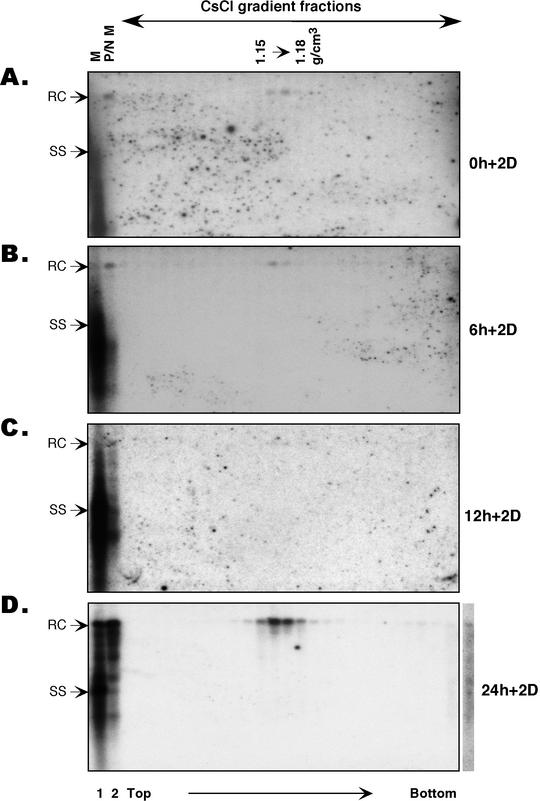
Intrinsic secretion competence of nucleocapsids of different maturity. Culture medium was collected from the synchronized replication system after viral DNA synthesis was allowed for 0, 6, 12, and 24 h and virion accumulation proceeded for the following 2 days (Fig. 2A); the medium samples are designated 0 h + 2 day (2D) (A), 6 h + 2 day (B), 12 h + 2 day (C), and 24 h + 2 day medium (D), respectively. Viral particles were pelleted from the medium by ultracentrifugation and were subjected to extensive proteolysis and nuclease digestion with protease and DNase I, followed by isopycnic CsCl gradient ultracentrifugation. The direction of sedimentation is indicated. Viral DNA was extracted from the untreatedmedium (lanes 1, M), the digested medium (lanes 2, P/N M), and the gradient fractions and was analyzed by Southern blotting. Sevenfold more medium was loaded in lane 2 than in lane 1. The density of the fractions containing authentic virions spanned 1.15 to 1.18 g/cm3. Note the absence of immature DNA species in the virion fractions at all time points. Residual naked nucleocapsids remaining after pronase-DNase I digestion migrated toward the bottom of the CsCl gradient, well separated from the virions (D). The extra lane on the right of panel D is a longer exposure of the last lane in the panel, showing the presence of immature viral DNA species at the bottom of the CsCl gradient. RC, RC DNA; ss, ss DNA. These experiments were repeated twice with essentially the same results obtained.
References
-
- Jilbert, A. R., D. S. Miller, C. A. Scougall, H. Turnbull, and C. J. Burrell. 1996. Kinetics of duck hepatitis B virus infection following low dose virus inoculation: one virus DNA genome is infectious in neonatal ducks. Virology 226:338-345. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
