

Tolerance for mutations and chemical modifications in a siRNA
- PMID: 12527766
- PMCID: PMC140512
- DOI: 10.1093/nar/gkg147
Tolerance for mutations and chemical modifications in a siRNA
Abstract
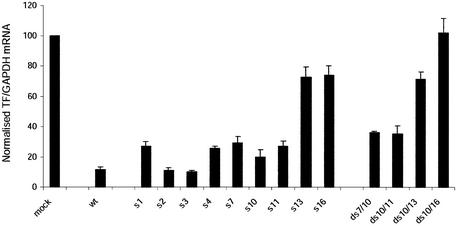
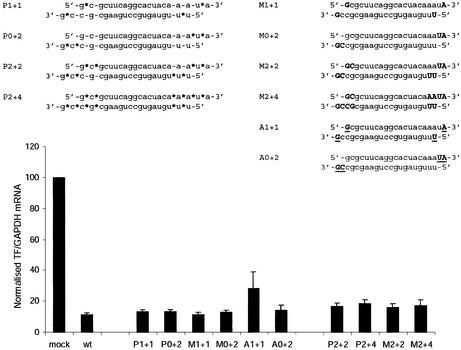
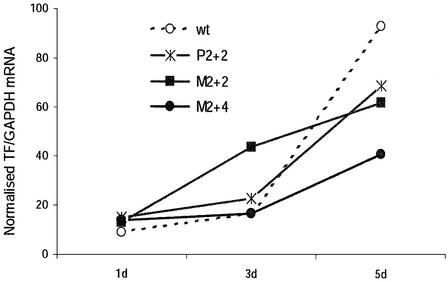
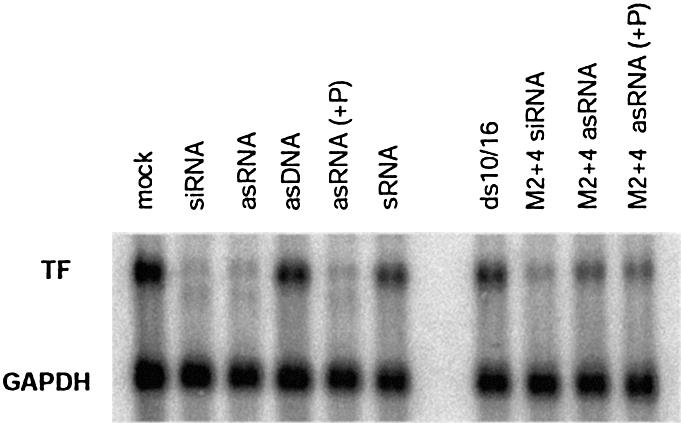
Short interfering RNA (siRNA), the active agent of RNA interference, shows promise of becoming a valuable tool in both basic and clinical research. We explore the tolerance to mutations and chemical modifications in various parts of the two 21-nt strands of a siRNA targeting the blood clotting initiator Tissue Factor. The mutations were G/C transversions. The chemical modifications were 2'-O-methylation, 2'-O-allylation and phosphorothioates. We found that siRNA generally tolerated mutations in the 5' end, while the 3' end exhibited low tolerance. This observation may facilitate the design of siRNA for specific targeting of transcripts containing single nucleotide polymorphisms. We further demonstrate that in our system the single antisense strand of the wild-type siRNA is almost as effective as the siRNA duplex, while the corresponding methylated M2+4 version of the antisense had reduced activity. Most of the chemically modified versions tested had near-wild-type initial activity, while the long-term activity was increased for certain siRNA species. Our results may improve the design of siRNAs for in vivo experiments.
Figures

References
-
- Fire A., Xu,S., Montgomery,M.K., Kostas,S.A., Driver,S.E. and Mello,C.C. (1998) Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature, 391, 806–811. - PubMed
-
- Elbashir S.M., Harborth,J., Lendeckel,W., Yalcin,A., Weber,K. and Tuschl,T. (2001) Duplexes of 21-nucleotide RNAs mediate RNA interference in cultured mammalian cells. Nature, 411, 494–498. - PubMed
-
- Lee N.S., Dohjima,T., Bauer,G., Li,H., Li,M.J., Ehsani,A., Salvaterra,P. and Rossi,J. (2002) Expression of small interfering RNAs targeted against HIV-1 rev transcripts in human cells. Nat. Biotechnol., 19, 500–505. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
