

Systematic mutagenesis of the Saccharomyces cerevisiae MLH1 gene reveals distinct roles for Mlh1p in meiotic crossing over and in vegetative and meiotic mismatch repair
- PMID: 12529393
- PMCID: PMC140715
- DOI: 10.1128/MCB.23.3.873-886.2003
Systematic mutagenesis of the Saccharomyces cerevisiae MLH1 gene reveals distinct roles for Mlh1p in meiotic crossing over and in vegetative and meiotic mismatch repair
Abstract
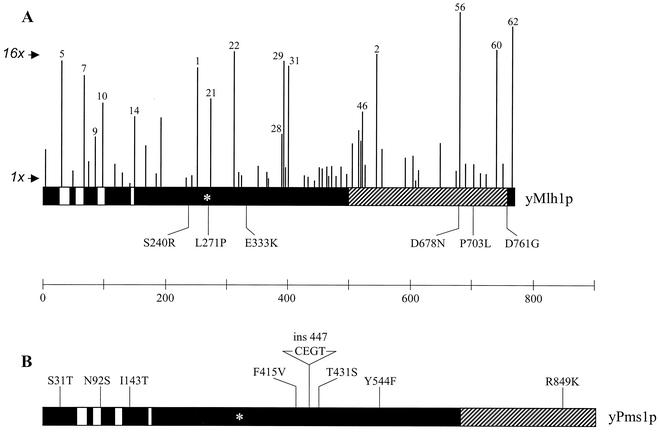
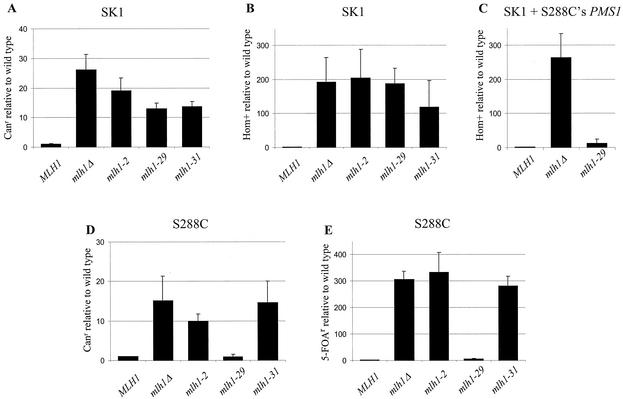
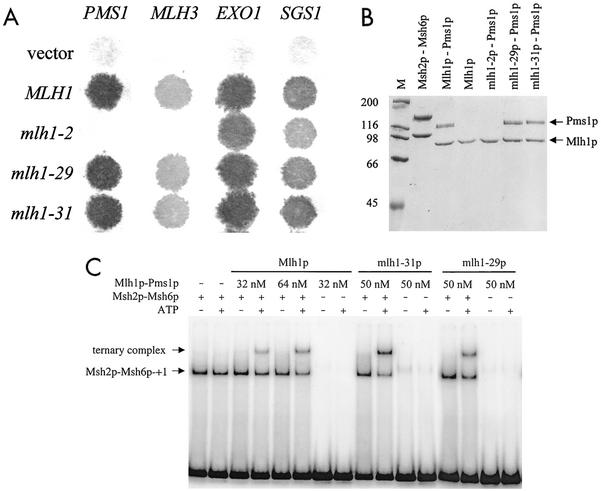
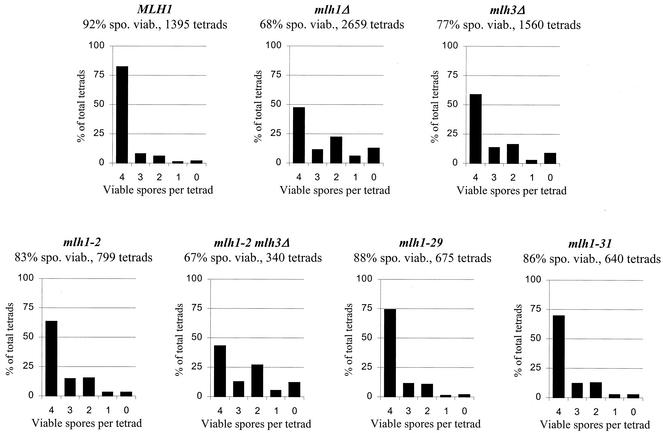
In eukaryotic cells, DNA mismatch repair is initiated by a conserved family of MutS (Msh) and MutL (Mlh) homolog proteins. Mlh1 is unique among Mlh proteins because it is required in mismatch repair and for wild-type levels of crossing over during meiosis. In this study, 60 new alleles of MLH1 were examined for defects in vegetative and meiotic mismatch repair as well as in meiotic crossing over. Four alleles predicted to disrupt the Mlh1p ATPase activity conferred defects in all functions assayed. Three mutations, mlh1-2, -29, and -31, caused defects in mismatch repair during vegetative growth but allowed nearly wild-type levels of meiotic crossing over and spore viability. Surprisingly, these mutants did not accumulate high levels of postmeiotic segregation at the ARG4 recombination hotspot. In biochemical assays, Pms1p failed to copurify with mlh1-2, and two-hybrid studies indicated that this allele did not interact with Pms1p and Mlh3p but maintained wild-type interactions with Exo1p and Sgs1p. mlh1-29 and mlh1-31 did not alter the ability of Mlh1p-Pms1p to form a ternary complex with a mismatch substrate and Msh2p-Msh6p, suggesting that the region mutated in these alleles could be responsible for signaling events that take place after ternary complex formation. These results indicate that mismatches formed during genetic recombination are processed differently than during replication and that, compared to mismatch repair functions, the meiotic crossing-over role of MLH1 appears to be more resistant to mutagenesis, perhaps indicating a structural role for Mlh1p during crossing over.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases