

Hydroxyurea induces fetal hemoglobin by the nitric oxide-dependent activation of soluble guanylyl cyclase
- PMID: 12531879
- PMCID: PMC151872
- DOI: 10.1172/JCI16672
Hydroxyurea induces fetal hemoglobin by the nitric oxide-dependent activation of soluble guanylyl cyclase
Abstract
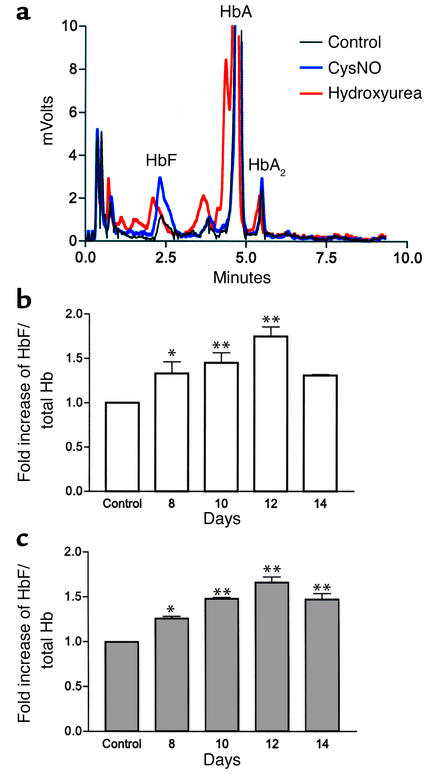
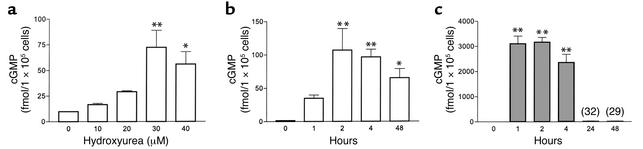
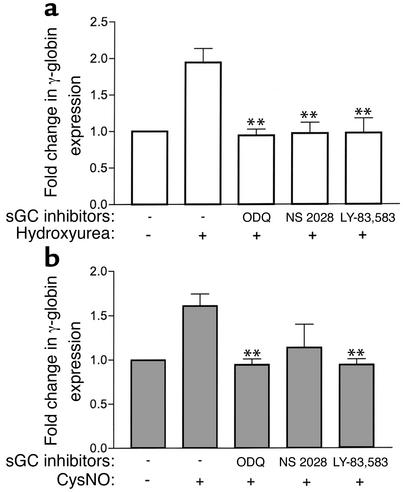
Hydroxyurea treatment of patients with sickle-cell disease increases fetal hemoglobin (HbF), which reduces hemoglobin S polymerization and clinical complications. Despite its use in the treatment of myeloproliferative diseases for over 30 years, its mechanism of action remains uncertain. Recent studies have demonstrated that hydroxyurea generates the nitric oxide (NO) radical in vivo, and we therefore hypothesized that NO-donor properties might determine the hemoglobin phenotype. We treated both K562 erythroleukemic cells and human erythroid progenitor cells with S-nitrosocysteine (CysNO), an NO donor, and found similar dose- and time-dependent induction of gamma-globin mRNA and HbF protein as we observed with hydroxyurea. Both hydroxyurea and CysNO increased cGMP levels, and the guanylyl cyclase inhibitors ODQ, NS 2028, and LY 83,538 abolished both the hydroxyurea- and CysNO-induced gamma-globin expression. These data provide strong evidence for an NO-derived mechanism for HbF induction by hydroxyurea and suggest possibilities for therapies based on NO-releasing or -potentiating agents.
Figures
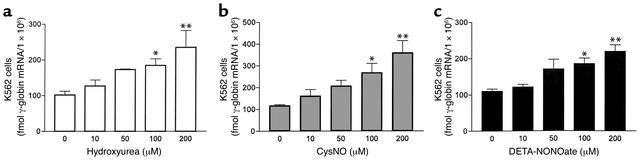
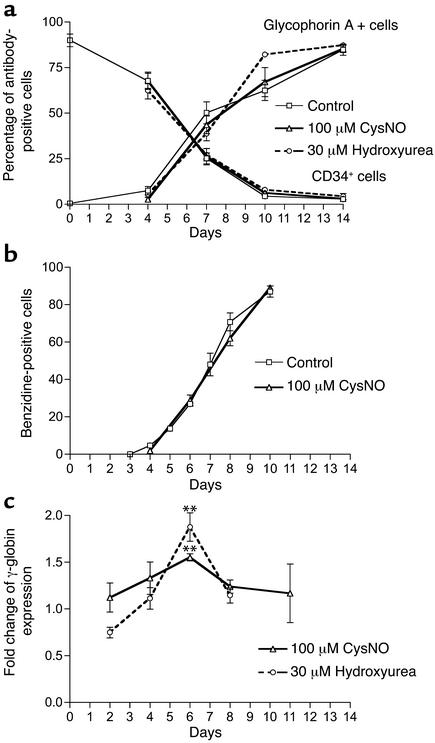
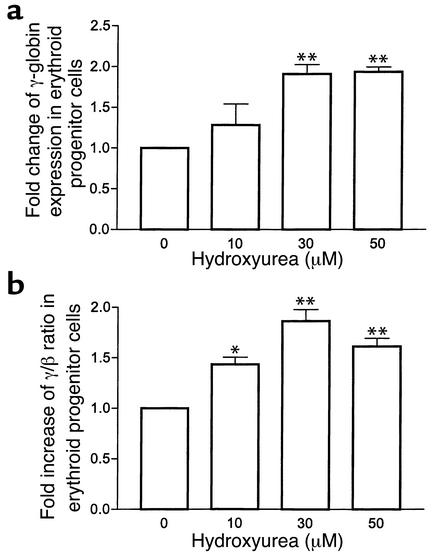
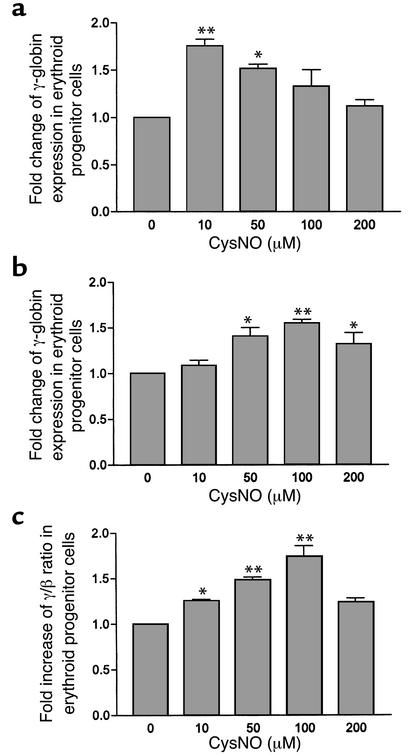
Comment on
-
A role for nitric oxide in hydroxyurea-mediated fetal hemoglobin induction.J Clin Invest. 2003 Jan;111(2):171-2. doi: 10.1172/JCI17597. J Clin Invest. 2003. PMID: 12531869 Free PMC article. No abstract available.
References
-
- Charache S, et al. Effect of hydroxyurea on the frequency of painful crises in sickle cell anemia. Investigators of the Multicenter Study of Hydroxyurea in Sickle Cell Anemia. N. Engl. J. Med. 1995;332:1317–1322. - PubMed
-
- Charache S. Mechanism of action of hydroxyurea in the management of sickle cell anemia in adults. Semin. Hematol. 1997;34:15–21. - PubMed
-
- Atweh GF, Schechter AN. Pharmacologic induction of fetal hemoglobin: raising the therapeutic bar in sickle cell disease. Curr. Opin. Hematol. 2001;8:123–130. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
