

Agrobacterium tumefaciens twin-arginine-dependent translocation is important for virulence, flagellation, and chemotaxis but not type IV secretion
- PMID: 12533451
- PMCID: PMC142831
- DOI: 10.1128/JB.185.3.760-771.2003
Agrobacterium tumefaciens twin-arginine-dependent translocation is important for virulence, flagellation, and chemotaxis but not type IV secretion
Abstract
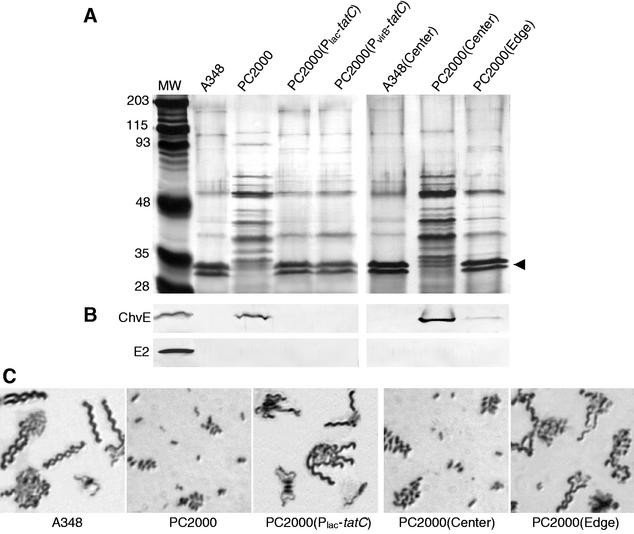
This study characterized the contribution of the twin-arginine translocation (TAT) pathway to growth, motility, and virulence of the phytopathogen Agrobacterium tumefaciens. In contrast to wild-type strain A348, a tatC null mutant failed to export the green fluorescent protein fused to the trimethylamine N-oxide reductase (TorA) signal sequence or to grow on nitrate as a sole electron acceptor during anaerobic growth. The tatC mutant displayed defects in growth rate and cell division but not in cell viability, and it also released abundant levels of several proteins into the culture supernatant when grown in rich medium or in vir induction minimal medium. Nearly all A348 cells were highly motile in both rich and minimal media. By contrast, approximately 0.1% of the tatC mutant cells were motile in rich medium, and <0.01% were motile in vir induction medium. Nonmotile tatC mutant cells lacked detectable flagella, whereas motile tatC mutant cells collected from the edge of a motility halo possessed flagella but not because of reversion to a functional TAT system. Motile tatC cells failed to exhibit chemotaxis toward sugars under aerobic conditions or towards nitrate under anaerobic conditions. The tatC mutant was highly attenuated for virulence, only occasionally (approximately 15% of inoculations) inciting formation of small tumors on plants after a prolonged incubation period of 6 to 8 weeks. However, an enriched subpopulation of motile tatC mutants exhibited enhanced virulence compared to the nonmotile variants. Finally, the tatC mutant transferred T-DNA and protein effectors to plant cells and a mobilizable IncQ plasmid to agrobacterial recipients at wild-type levels. Together, our findings establish that, in addition to its role in secretion of folded cofactor-bound enzymes functioning in alternative respiration, the TAT system of A. tumefaciens is an important virulence determinant. Furthermore, this secretion pathway contributes to flagellar biogenesis and chemotactic responses but not to sensory perception of plant signals or the assembly of a type IV secretion system.
Figures





References
-
- Alvarez-Martinez, M. T., J. Machold, C. Weise, H. Schmidt-Eisenlohr, C. Baron, and B. Rouot. 2001. The Brucella suis homologue of the Agrobacterium tumefaciens chromosomal virulence operon chvE is essential for sugar utilization but not for survival in macrophages. J. Bacteriol. 183:5343-5351. - PMC - PubMed
-
- Angelini, S., R. Moreno, K. Gouffi, C. Santini, A. Yamagishi, J. Berenguer, and L. Wu. 2001. Export of Thermus thermophilus alkaline phosphatase via the twin-arginine translocation pathway in Escherichia coli. FEBS Lett. 506:103-107. - PubMed
-
- Bagdasarian, M., R. Lurz, B. Ruckert, F. C. Franklin, M. M. Bagdasarian, J. Frey, and K. N. Timmis. 1981. Specific-purpose plasmid cloning vectors. II. Broad host range, high copy number, RSF1010-derived vectors, and a host-vector system for gene cloning in Pseudomonas. Gene 16:237-247. - PubMed
-
- Bash, R., and A. G. Matthysse. 2002. Attachment to roots and virulence of a chvB mutant of Agrobacterium tumefaciens are temperature sensitive. Mol. Plant-Microbe Interact. 15:160-163. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
