

A SecE mutation that modulates SecY-SecE translocase assembly, identified as a specific suppressor of SecY defects
- PMID: 12533470
- PMCID: PMC142837
- DOI: 10.1128/JB.185.3.948-956.2003
A SecE mutation that modulates SecY-SecE translocase assembly, identified as a specific suppressor of SecY defects
Abstract
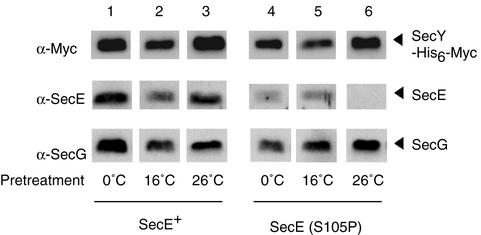
The SecY39(Cs) (cold-sensitive) alteration of Arg357 results in a defect of translocation initiation. As a means to dissect the Sec translocation machinery, we isolated mutations that act as suppressors of the secY39 defect. A specific secE mutation, designated secE105, was thus isolated. This mutation proved to be identical with the prlG2 mutation and to suppress a number of cold-sensitive secY mutations. However, other prlG mutations did not effectively suppress the secY defects. Evidence indicates that the Ser105-to-Pro alteration in the C-terminal transmembrane segment of SecE weakens SecY-SecE association. In vitro analyses showed that the SecE(S105P) alteration preferentially stimulates the initial phase of translocation. It is suggested that the S105P alteration affects the SecYEG channel such that it is more prone to open and to accept the translocation initiation domain of a preprotein molecule.
Figures






Similar articles
-
Preprotein translocation by a hybrid translocase composed of Escherichia coli and Bacillus subtilis subunits.J Bacteriol. 1999 Nov;181(22):7021-7. doi: 10.1128/JB.181.22.7021-7027.1999. J Bacteriol. 1999. PMID: 10559168 Free PMC article.
-
Evaluating the oligomeric state of SecYEG in preprotein translocase.EMBO J. 2000 Aug 15;19(16):4393-401. doi: 10.1093/emboj/19.16.4393. EMBO J. 2000. PMID: 10944122 Free PMC article.
-
Characterization of the supporting role of SecE in protein translocation.FEBS Lett. 2013 Sep 17;587(18):3083-8. doi: 10.1016/j.febslet.2013.07.046. Epub 2013 Aug 13. FEBS Lett. 2013. PMID: 23954289
-
The bacterial Sec-translocase: structure and mechanism.Philos Trans R Soc Lond B Biol Sci. 2012 Apr 19;367(1592):1016-28. doi: 10.1098/rstb.2011.0201. Philos Trans R Soc Lond B Biol Sci. 2012. PMID: 22411975 Free PMC article. Review.
-
The Sec system.Curr Opin Microbiol. 1998 Apr;1(2):216-22. doi: 10.1016/s1369-5274(98)80014-3. Curr Opin Microbiol. 1998. PMID: 10066476 Review.
Cited by
-
Mutational analysis of transmembrane regions 3 and 4 of SecY, a central component of protein translocase.J Bacteriol. 2004 Jun;186(12):3960-9. doi: 10.1128/JB.186.12.3960-3969.2004. J Bacteriol. 2004. PMID: 15175310 Free PMC article.
-
Purification, crystallization and preliminary X-ray diffraction of SecDF, a translocon-associated membrane protein, from Thermus thermophilus.Acta Crystallogr Sect F Struct Biol Cryst Commun. 2006 Apr 1;62(Pt 4):376-80. doi: 10.1107/S1744309106007779. Epub 2006 Mar 25. Acta Crystallogr Sect F Struct Biol Cryst Commun. 2006. PMID: 16582489 Free PMC article.
-
Identification and characterization of a translation arrest motif in VemP by systematic mutational analysis.J Biol Chem. 2018 Feb 23;293(8):2915-2926. doi: 10.1074/jbc.M117.816561. Epub 2018 Jan 9. J Biol Chem. 2018. PMID: 29317498 Free PMC article.
-
Dynamics of SecY translocons with translocation-defective mutations.Structure. 2010 Jul 14;18(7):847-57. doi: 10.1016/j.str.2010.04.010. Structure. 2010. PMID: 20637421 Free PMC article.
-
Different modes of SecY-SecA interactions revealed by site-directed in vivo photo-cross-linking.Proc Natl Acad Sci U S A. 2006 Oct 31;103(44):16159-64. doi: 10.1073/pnas.0606390103. Epub 2006 Oct 23. Proc Natl Acad Sci U S A. 2006. PMID: 17060619 Free PMC article.
References
-
- Akiyama, Y., and K. Ito. 1990. SecY protein, a membrane embedded secretion factor of E. coli, is cleaved by the OmpT protease in vitro. Biochem. Biophys. Res. Commun. 167:711-715. - PubMed
-
- Akiyama, Y., T. Ogura, and K. Ito. 1994. Involvement of FtsH in protein assembly into and through the membrane. J. Biol. Chem. 269:5218-5224. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
