

Extracellular Ca2+ depletion contributes to fast activity-dependent modulation of synaptic transmission in the brain
- PMID: 12546823
- PMCID: PMC3375894
- DOI: 10.1016/s0896-6273(03)00025-4
Extracellular Ca2+ depletion contributes to fast activity-dependent modulation of synaptic transmission in the brain
Abstract
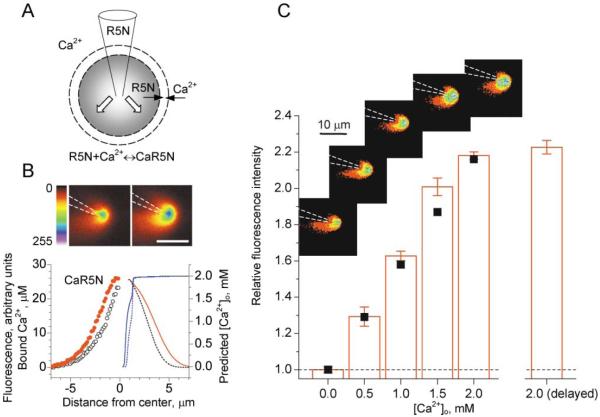
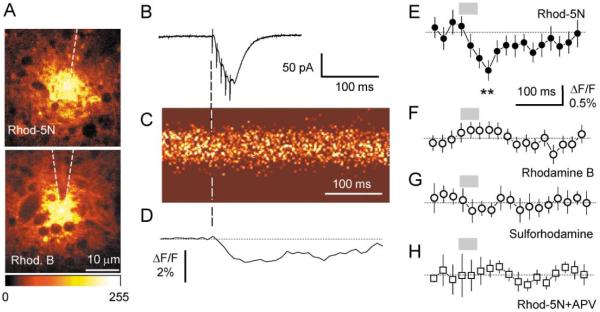
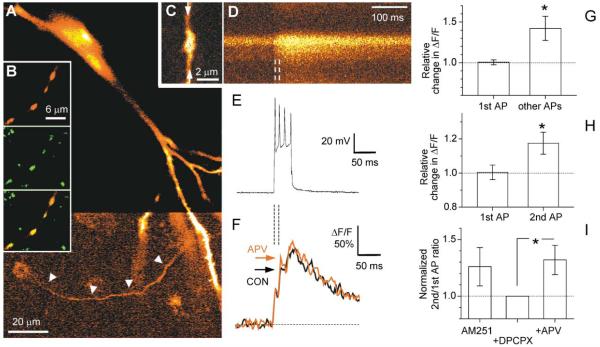
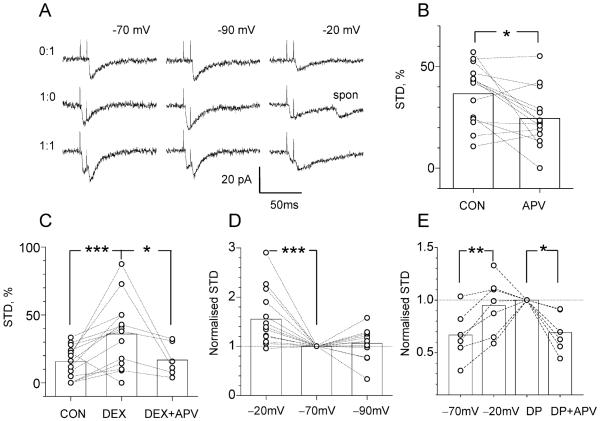
Synaptic activation is associated with rapid changes in intracellular Ca(2+), while the extracellular Ca(2+) level is generally assumed to be constant. Here, using a novel optical method to measure changes in extracellular Ca(2+) at high spatial and temporal resolution, we find that brief trains of synaptic transmission in hippocampal area CA1 induce transient depletion of extracellular Ca(2+). We show that this depletion, which depends on postsynaptic NMDA receptor activation, decreases the Ca(2+) available to enter individual presynaptic boutons of CA3 pyramidal cells. This in turn reduces the probability of consecutive synaptic releases at CA3-CA1 synapses and therefore contributes to short-term paired-pulse depression of minimal responses. This activity-dependent depletion of extracellular Ca(2+) represents a novel form of fast retrograde synaptic signaling that can modulate glutamatergic information transfer in the brain.
Figures
References
-
- Bellingham MC, Walmsley B. A novel presynaptic inhibitory mechanism underlies paired pulse depression at a fast central synapse. Neuron. 1999;23:159–170. - PubMed
-
- Berretta N, Rossokhin AV, Kasyanov AM, Sokolov MV, Cherubini E, Voronin LL. Postsynaptic hyperpolarization increases the strength of AMPA-mediated synaptic transmission at large synapses between mossy fibers and CA3 pyramidal cells. Neuropharmacology. 2000;39:2288–2301. - PubMed
-
- Brown EM, Vassiliev PM, Hebert SC. Calcium ions as extracellular messengers. Cell. 1995;83:679–682. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
