

Early induction and maintenance of Env-specific T-helper cells following human immunodeficiency virus type 1 infection
- PMID: 12552005
- PMCID: PMC141076
- DOI: 10.1128/jvi.77.4.2663-2674.2003
Early induction and maintenance of Env-specific T-helper cells following human immunodeficiency virus type 1 infection
Abstract
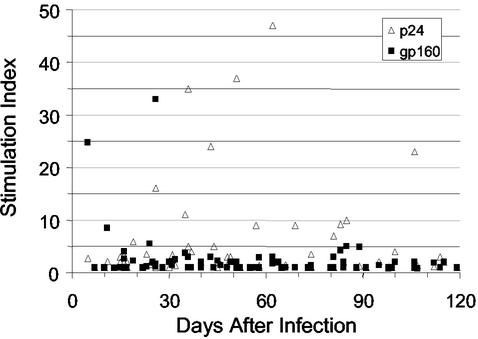
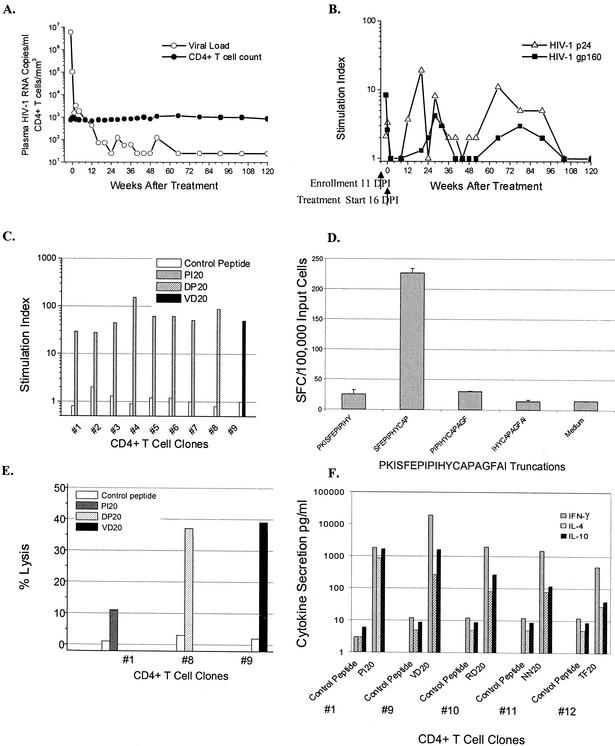
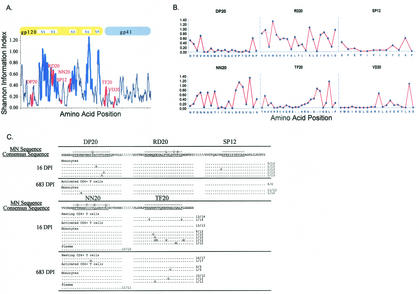
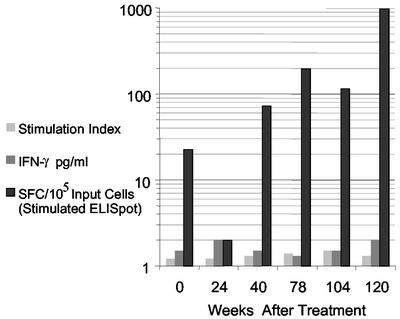
Mounting evidence points to a role for CD4+ T-helper (Th) cell activities in controlling human immunodeficiency virus type 1 (HIV-1) infection. To determine the induction and evolution of Th responses following acute infection, we prospectively analyzed Env- and Gag-specific Th responses longitudinally for 92 patients with acute (n = 28) or early (n = 64) HIV-1 infection (median, 55 days postinfection [DPI]). The probability of detecting HIV-1-specific lymphoproliferative responses was remarkably low, and when present, the responses were more likely to be Gag specific than Env specific (16 versus 5%). Env-specific responses were significantly more common in patients presenting at <30 DPI than in those presenting at 30 to 365 DPI (21 versus 0.5%, P = 0.001). By contrast, Gag-specific responses occurred with similar frequencies among subjects presenting at <30 DPI and 30 to 365 DPI (13 versus 17%, P = 0.6). After treatment, and regardless of the duration of infection before therapy, Gag-specific Th responses predominated. Furthermore, some acutely infected subjects lost detectable Env-specific Th proliferative responses, which failed to reemerge upon treatment. Detailed analysis for one such subject revealed Env-specific lymphoproliferation at 11 DPI but no detectable Env-specific lymphoproliferation or ex vivo gamma interferon (IFN-gamma) secretion at multiple subsequent time points. Env-specific CD4+ T-cell clones from 11 DPI recognized six epitopes in both conserved and variable regions within gp120 and gp41, exhibited major histocompatibility complex-restricted cytotoxicity, and secreted high levels of antiviral cytokines. T-cell receptor clonal transcript analyses and autologous virus sequencing revealed that Th cells induced during acute infection were maintained and there were no Th escape mutations. Subsequent analysis for this subject and six of seven others revealed detectable IFN-gamma-secreting cells, but only following in vitro gp160 stimulation. In summary, we conclude that Env-specific Th responses are elicited very early in acute infection and may precede Gag-specific responses. The inability to detect Env-specific Th responses over time and despite antiretroviral therapy may reflect low frequencies and impaired proliferative capacity, and viral escape is not necessary for this to occur.
Figures
Similar articles
-
Reconstitution of virus-specific CD4 proliferative responses in pediatric HIV-1 infection.J Immunol. 2003 Dec 15;171(12):6968-75. doi: 10.4049/jimmunol.171.12.6968. J Immunol. 2003. PMID: 14662905
-
Role of human immunodeficiency virus (HIV)-specific T-cell immunity in control of dual HIV-1 and HIV-2 infection.J Virol. 2007 Sep;81(17):9061-71. doi: 10.1128/JVI.00117-07. Epub 2007 Jun 20. J Virol. 2007. PMID: 17582003 Free PMC article.
-
Comparison of human immunodeficiency virus (HIV)-specific T-cell responses in HIV-1- and HIV-2-infected individuals in Senegal.J Virol. 2004 Dec;78(24):13934-42. doi: 10.1128/JVI.78.24.13934-13942.2004. J Virol. 2004. PMID: 15564501 Free PMC article.
-
Cellular immune response to human immunodeficiency virus.AIDS. 2001 Feb;15 Suppl 2:S16-21. doi: 10.1097/00002030-200102002-00004. AIDS. 2001. PMID: 11424972 Review.
-
Cytokines in immune regulation/pathogenesis in HIV infection.Ciba Found Symp. 1995;195:142-7; discussion 147-53. doi: 10.1002/9780470514849.ch10. Ciba Found Symp. 1995. PMID: 8724835 Review.
Cited by
-
Major histocompatibility complex class II molecule-human immunodeficiency virus peptide analysis using a microarray chip.Clin Vaccine Immunol. 2009 Apr;16(4):567-73. doi: 10.1128/CVI.00441-08. Epub 2009 Feb 18. Clin Vaccine Immunol. 2009. PMID: 19225081 Free PMC article.
-
A novel liposome-based adjuvant CAF01 for induction of CD8(+) cytotoxic T-lymphocytes (CTL) to HIV-1 minimal CTL peptides in HLA-A*0201 transgenic mice.PLoS One. 2009 Sep 11;4(9):e6950. doi: 10.1371/journal.pone.0006950. PLoS One. 2009. PMID: 19759892 Free PMC article.
-
Progress in defining CD4 helper cell responses in chronic viral infections.J Exp Med. 2003 Dec 15;198(12):1773-7. doi: 10.1084/jem.20031947. J Exp Med. 2003. PMID: 14676292 Free PMC article. No abstract available.
-
Antigen stimulation induces HIV envelope gp120-specific CD4(+) T cells to secrete CCR5 ligands and suppress HIV infection.Virology. 2007 Dec 5;369(1):214-25. doi: 10.1016/j.virol.2007.07.031. Epub 2007 Sep 4. Virology. 2007. PMID: 17765942 Free PMC article.
-
Functional properties and epitope characteristics of T-cells recognizing natural HIV-1 variants.Vaccine. 2009 Nov 12;27(48):6678-87. doi: 10.1016/j.vaccine.2009.08.093. Epub 2009 Sep 9. Vaccine. 2009. PMID: 19747576 Free PMC article.
References
-
- Akatsuka, Y., E. G. Martin, A. Madonik, A. A. Barsoukov, and J. A. Hansen. 1999. Rapid screening of T-cell receptor (TCR) variable gene usage by multiplex PCR: application for assessment of clonal composition. Tissue Antigens 53:122-134. - PubMed
-
- Berzofsky, J. A., A. Bensussan, K. B. Cease, J. F. Bourge, R. Cheynier, Z. Lurhuma, J. J. Salaun, R. C. Gallo, G. M. Shearer, and D. Zagury. 1988. Antigenic peptides recognized by T lymphocytes from AIDS viral envelope-immune humans. Nature 334:706-708. - PubMed
-
- Binley, J. M., D. S. Schiller, G. M. Ortiz, A. Hurley, D. F. Nixon, M. M. Markowitz, and J. P. Moore. 2000. The relationship between T cell proliferative responses and plasma viremia during treatment of human immunodeficiency virus type 1 infection with combination antiretroviral therapy. J. Infect. Dis. 181:1249-1263. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
