

High mutant frequency in populations of a DNA virus allows evasion from antibody therapy in an immunodeficient host
- PMID: 12552010
- PMCID: PMC141124
- DOI: 10.1128/jvi.77.4.2701-2708.2003
High mutant frequency in populations of a DNA virus allows evasion from antibody therapy in an immunodeficient host
Abstract
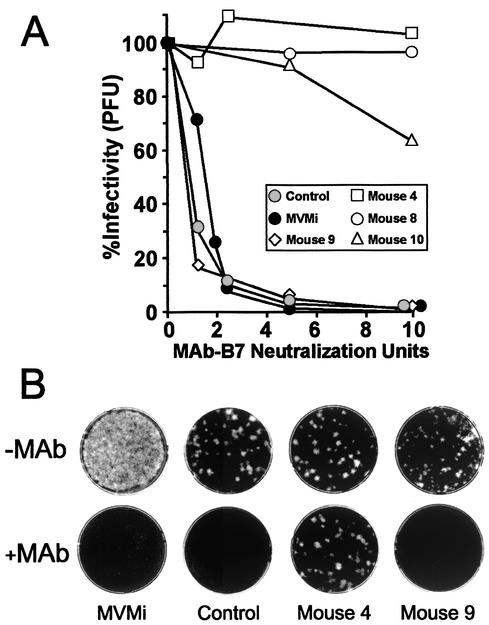
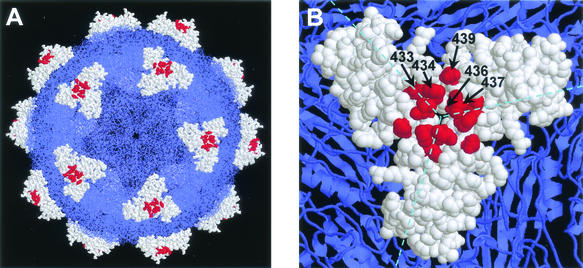
The degree of genetic heterogeneity of DNA virus populations in nature and its consequences for disease control are virtually unknown. The parvovirus minute virus of mice (MVMi) was used here to investigate (i) the frequency of antibody-escape mutants in populations of a DNA virus and (ii) the ability of a DNA virus to evade in the long-term a passive monoclonal antibody (MAb) therapy in an immunodeficient natural host. Independent clonal populations of MVMi harbored a high proportion of mutants resistant to neutralizing MAb (mutant frequency = [2.8 +/- 0.5] x 10(-5)) that rapidly evolved under antibody pressure in culture to become mixtures dominated by genotypically diverse escape mutants. Immunodeficient mice naturally infected with clonal populations of MVMi and subsequently treated by intravenous injections of MAb were initially protected from the characteristic viral induced lethal leukopenia. However, some treated animals developed a delayed severe leukopenic syndrome associated with the emergence of genetically heterogeneous populations of MAb-resistant mutants in the MVMi main target organs. The 11 plaque-purified viruses analyzed from an antibody-resistant population obtained from one animal corresponded to four different mutant genotypes, although their consensus sequence remained wild type. All cloned escape mutants harbored single radical amino acid changes within a stretch of seven residues in a surface-exposed loop at the threefold axes of the capsid. This antigenic site, which can tolerate radical changes preserving MVMi pathogenic potential, may thereby allow the virus to evade the immune control. These findings indicate a high genetic heterogeneity and rapid adaptation of populations of a mammal DNA virus in vivo and provide a genetic basis for the failure of passive immunotherapy in the natural host.
Figures
Similar articles
-
Evolution to pathogenicity of the parvovirus minute virus of mice in immunodeficient mice involves genetic heterogeneity at the capsid domain that determines tropism.J Virol. 2008 Feb;82(3):1195-203. doi: 10.1128/JVI.01692-07. Epub 2007 Nov 28. J Virol. 2008. PMID: 18045943 Free PMC article.
-
Enhanced cytoplasmic sequestration of the nuclear export receptor CRM1 by NS2 mutations developed in the host regulates parvovirus fitness.J Virol. 2004 Oct;78(19):10674-84. doi: 10.1128/JVI.78.19.10674-10684.2004. J Virol. 2004. PMID: 15367634 Free PMC article.
-
Parvovirus infection suppresses long-term repopulating hematopoietic stem cells.J Virol. 2003 Aug;77(15):8495-503. doi: 10.1128/jvi.77.15.8495-8503.2003. J Virol. 2003. PMID: 12857918 Free PMC article.
-
Parvovirus variation for disease: a difference with RNA viruses?Curr Top Microbiol Immunol. 2006;299:349-70. doi: 10.1007/3-540-26397-7_13. Curr Top Microbiol Immunol. 2006. PMID: 16568906 Review.
-
Minute virus of mice, a parvovirus, in complex with the Fab fragment of a neutralizing monoclonal antibody.J Virol. 2007 Sep;81(18):9851-8. doi: 10.1128/JVI.00775-07. Epub 2007 Jul 11. J Virol. 2007. PMID: 17626084 Free PMC article.
Cited by
-
In vitro and in vivo fitness of respiratory syncytial virus monoclonal antibody escape mutants.J Virol. 2006 Dec;80(23):11651-7. doi: 10.1128/JVI.01387-06. Epub 2006 Sep 27. J Virol. 2006. PMID: 17005645 Free PMC article.
-
Nuclear export of the nonenveloped parvovirus virion is directed by an unordered protein signal exposed on the capsid surface.J Virol. 2004 Oct;78(19):10685-94. doi: 10.1128/JVI.78.19.10685-10694.2004. J Virol. 2004. PMID: 15367635 Free PMC article.
-
A supraphysiological nuclear export signal is required for parvovirus nuclear export.Mol Biol Cell. 2008 Jun;19(6):2544-52. doi: 10.1091/mbc.e08-01-0009. Epub 2008 Apr 2. Mol Biol Cell. 2008. PMID: 18385513 Free PMC article.
-
Evolution to pathogenicity of the parvovirus minute virus of mice in immunodeficient mice involves genetic heterogeneity at the capsid domain that determines tropism.J Virol. 2008 Feb;82(3):1195-203. doi: 10.1128/JVI.01692-07. Epub 2007 Nov 28. J Virol. 2008. PMID: 18045943 Free PMC article.
-
Host-selected amino acid changes at the sialic acid binding pocket of the parvovirus capsid modulate cell binding affinity and determine virulence.J Virol. 2006 Feb;80(3):1563-73. doi: 10.1128/JVI.80.3.1563-1573.2006. J Virol. 2006. PMID: 16415031 Free PMC article.
References
-
- Agbandje-McKenna, M., A. LLamas-Saiz, F. Wang, P. Tattersall, and M. G. Rossmann. 1998. Functional implications of the structure of the murine parvovirus minute virus of mice. Structure 6:1369-1381. - PubMed
-
- Bloom, M. E., S. M. Best, S. F. Hayes, R. D. Wells, J. B. Wolfinbarger, R. McKenna, and M. Agbandje-McKenna. 2001. Identification of Aleutian mink disease parvovirus capsid sequences mediating antibody-dependent enhancement of infection, virus neutralization, and inmune complex formation. J. Virol. 75:11116-11127. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
