

Small ubiquitin-related modifier-1 modification mediates resolution of CREB-dependent responses to hypoxia
- PMID: 12552083
- PMCID: PMC298713
- DOI: 10.1073/pnas.0337412100
Small ubiquitin-related modifier-1 modification mediates resolution of CREB-dependent responses to hypoxia
Abstract
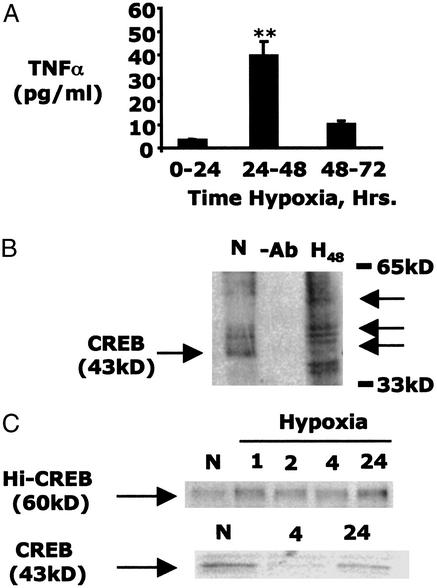
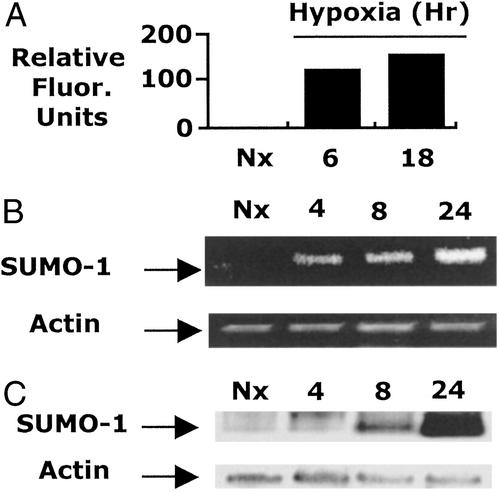
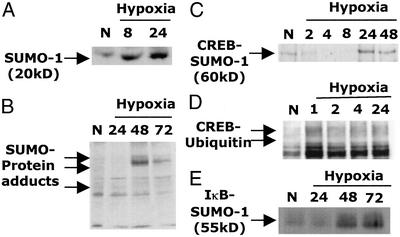
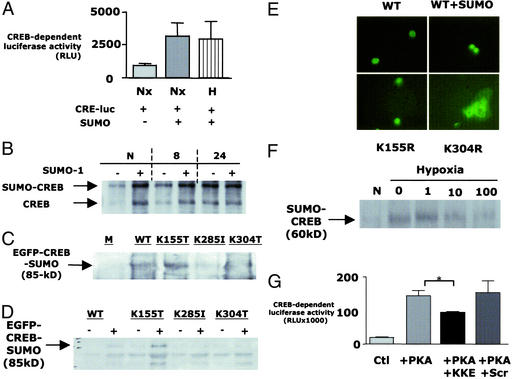
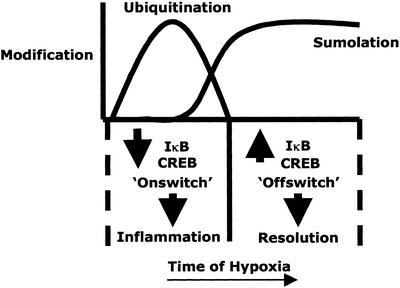
Phosphorylation-dependent ubiquitination combined with proteasomal degradation of transcriptional regulators is a recently appreciated mechanism for control of a number of inflammatory genes. Far less is known about the counterregulatory mechanisms that repress transcriptional activity in these pathways during resolution. Here, we investigated the transient nature of hypoxia-induced tumor necrosis factor (TNF)alpha in T84 cells, a process we have previously shown to involve phosphorylation-dependent degradation of the cAMP-response element-binding protein (CREB). Initial studies indicate hypoxia-induced TNFalpha to be a transient event, the resolution of which is associated with the appearance of a higher molecular weight modified form of CREB. Gene array analysis of mRNA derived from hypoxic cells identified a time-dependent induction of small ubiquitin-related modifier (SUMO)-1 mRNA. In prolonged hypoxia, CREB is posttranslationally modified by SUMO-1. Furthermore, SUMO-1 overexpression stabilizes CREB in hypoxia and enhances CREB-dependent reporter gene activity. Site-directed mutagenesis of lysine residues K285 and K304 identifies them as SUMO acceptors in vivo and in vitro. Mutation of K304 also results in loss of CREB nuclear localization, implying a role for SUMO-1 modification at this site in the subcellular localization of CREB. Thus, in prolonged hypoxia, CREB is modified by association with SUMO-1. Furthermore, we hypothesize that such an event stabilizes and promotes nuclear localization of CREB and thus complements an endogenous resolution phase for hypoxia-induced inflammatory processes.
Figures
References
-
- Taylor C T, Colgan S P. Pharmacol Res. 1999;16:1498–1505. - PubMed
-
- Bunn H F, Poyton R O. Physiol Rev. 1996;76:839–885. - PubMed
-
- Taylor C T, Dzus A L, Colgan S P. Gastroenterology. 1998;114:657–668. - PubMed
-
- Taylor C T, Fueki N, Agah A, Hershberg R M, Colgan S P. J Biol Chem. 1999;274:19447–19454. - PubMed
-
- Karin M, Ben-Neriah Y. Ann Rev Immunol. 2000;18:621–663. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
