

C-terminal domains implicated in the functional surface expression of potassium channels
- PMID: 12554641
- PMCID: PMC140720
- DOI: 10.1093/emboj/cdg035
C-terminal domains implicated in the functional surface expression of potassium channels
Abstract
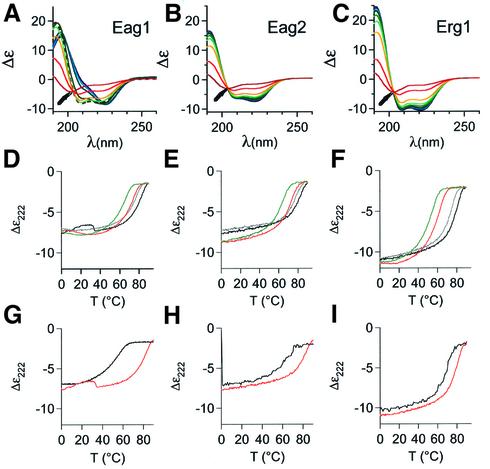
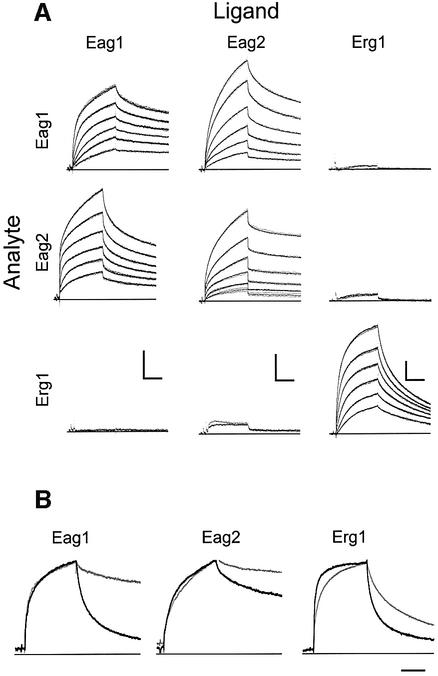
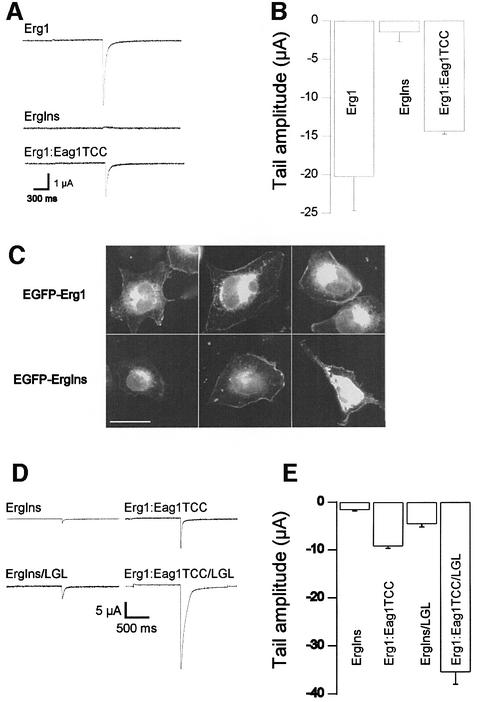
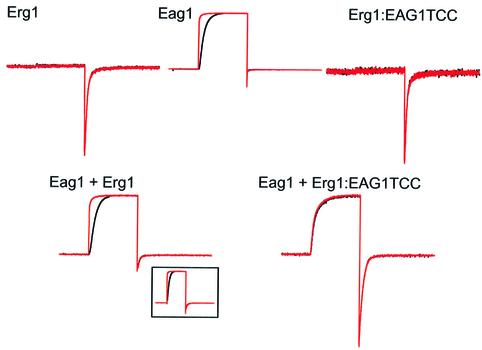
A short C-terminal domain is required for correct tetrameric assembly in some potassium channels. Here, we show that this domain forms a coiled coil that determines not only the stability but also the selectivity of the multimerization. Synthetic peptides comprising the sequence of this domain in Eag1 and other channels are able to form highly stable tetrameric coiled coils and display selective heteromultimeric interactions. We show that loss of function caused by disruption of this domain in Herg1 can be rescued by introducing the equivalent domain from Eag1, and that this chimeric protein can form heteromultimers with Eag1 while wild-type Erg1 cannot. Additionally, a short endoplasmic reticulum retention sequence closely preceding the coiled coil plays a crucial role for surface expression. Both domains appear to co-operate to form fully functional channels on the cell surface and are a frequent finding in ion channels. Many pathological phenotypes may be attributed to mutations affecting one or both domains.
Figures



References
-
- Biervert C., Schroeder,B.C., Kubisch,C., Berkovic,S.F., Propping,P., Jentsch,T.J. and Steinlein,O.K. (1998) A potassium channel mutation in neonatal human epilepsy. Science, 279, 403–406. - PubMed
-
- Bohm G., Muhr,R. and Jaenicke,R. (1992) Quantitative analysis of protein far UV circular dichroism spectra by neural networks. Protein Eng., 5, 191–195. - PubMed
-
- Brüggemann A., Pardo,L.A., Stühmer,W. and Pongs,O. (1993) Ether-à-go-go encodes a voltage-gated channel permeable to K+ and Ca2+ and modulated by cAMP. Nature 365, 445–448. - PubMed
-
- Cabezon E., Arechaga,I., Butler,P.J.G. and Walker,J.E. (2000) Dimerization of bovine F-1-ATPase by binding the inhibitor protein, IF1. J. Biol. Chem., 275, 28353–28355. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
