

Retromer and the sorting nexins Snx4/41/42 mediate distinct retrieval pathways from yeast endosomes
- PMID: 12554655
- PMCID: PMC140746
- DOI: 10.1093/emboj/cdg062
Retromer and the sorting nexins Snx4/41/42 mediate distinct retrieval pathways from yeast endosomes
Abstract
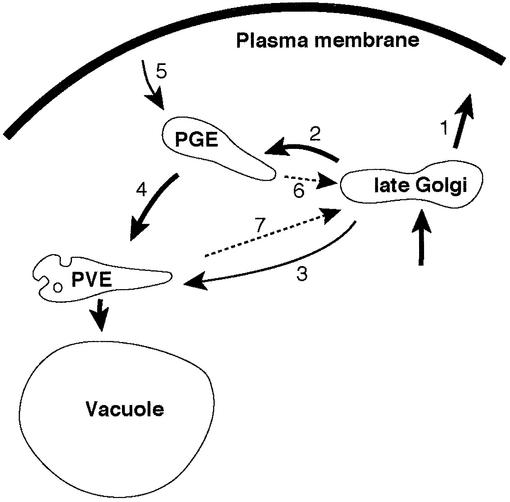
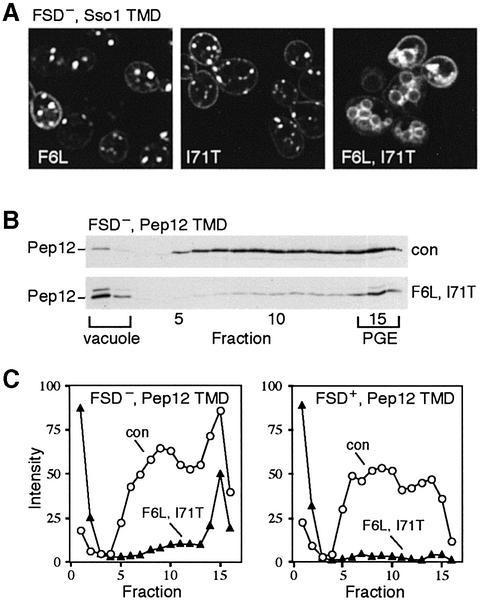
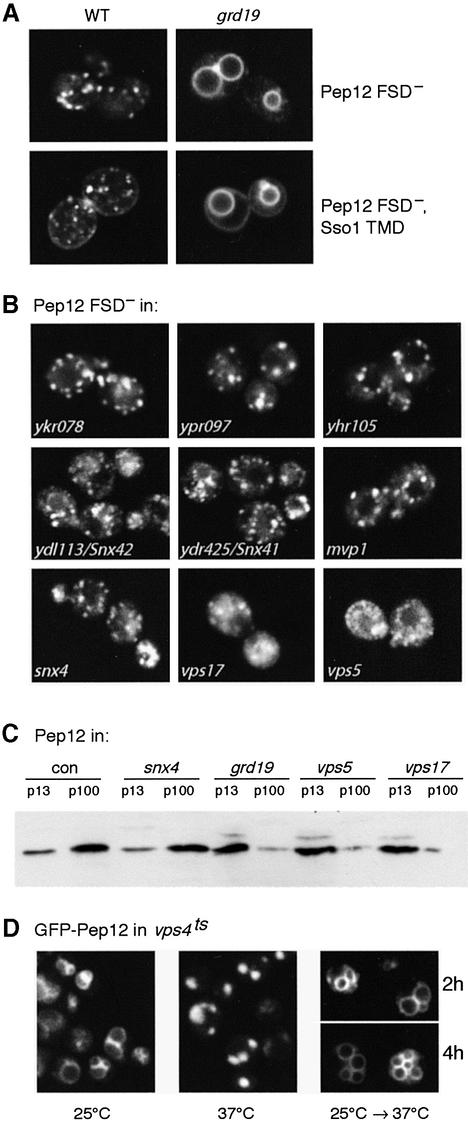
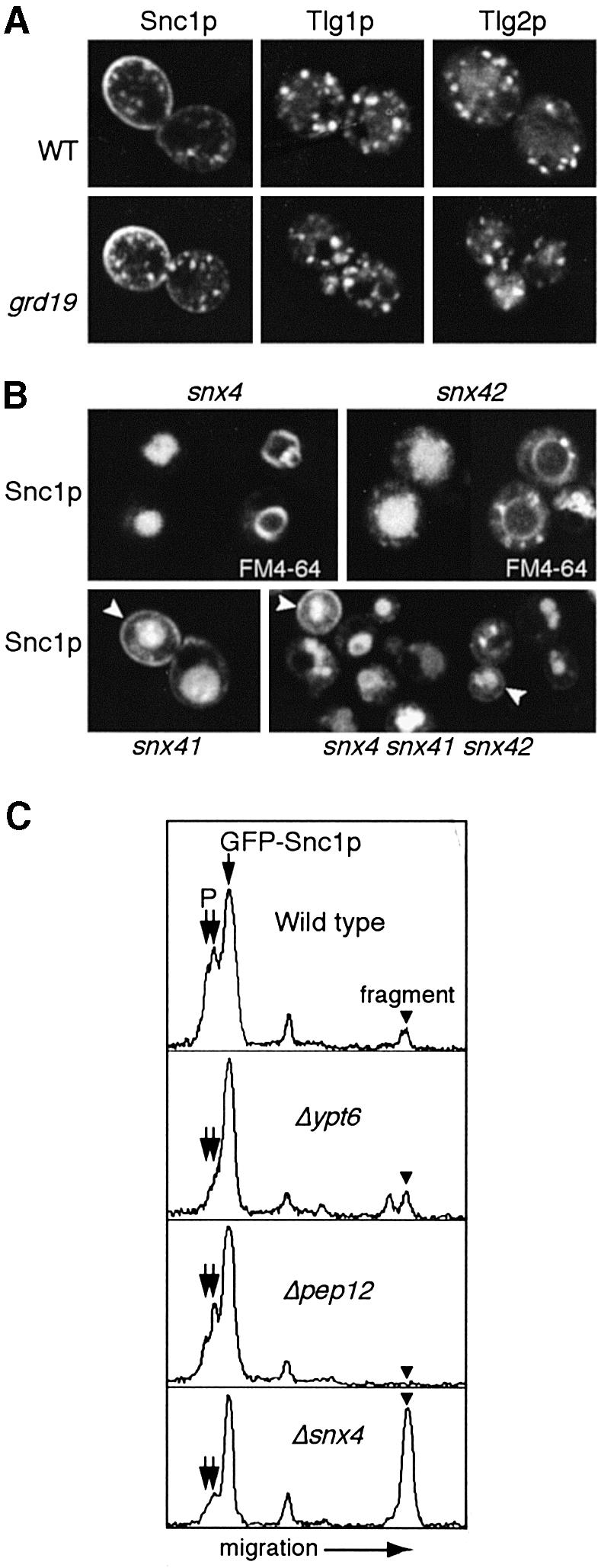
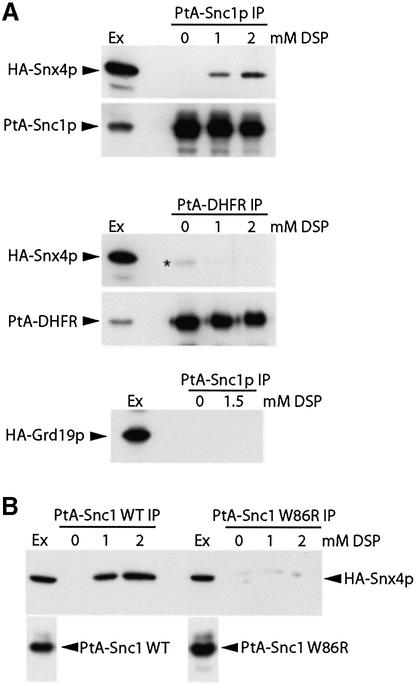
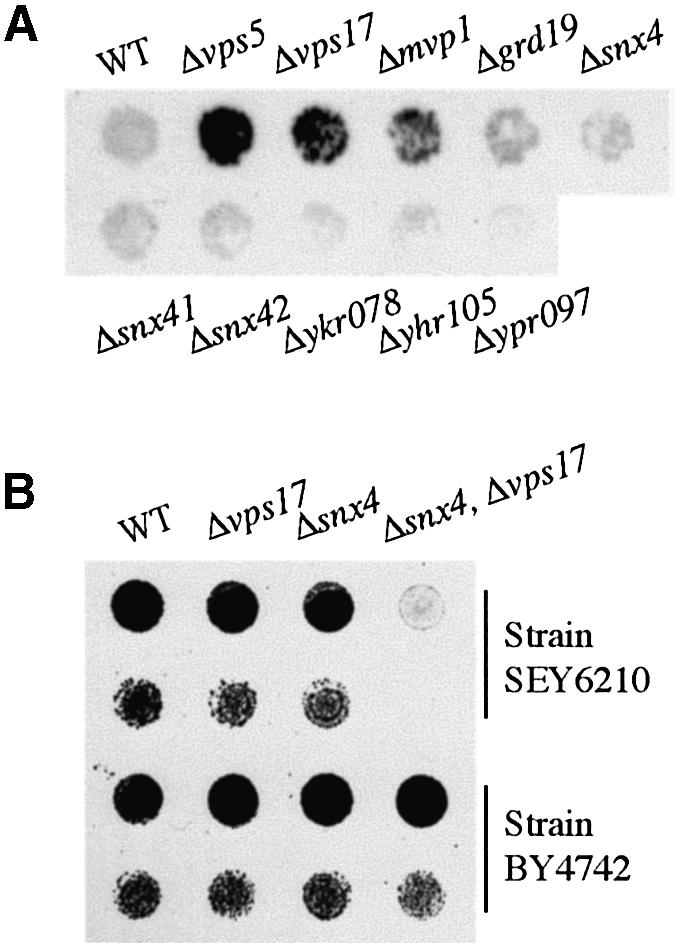
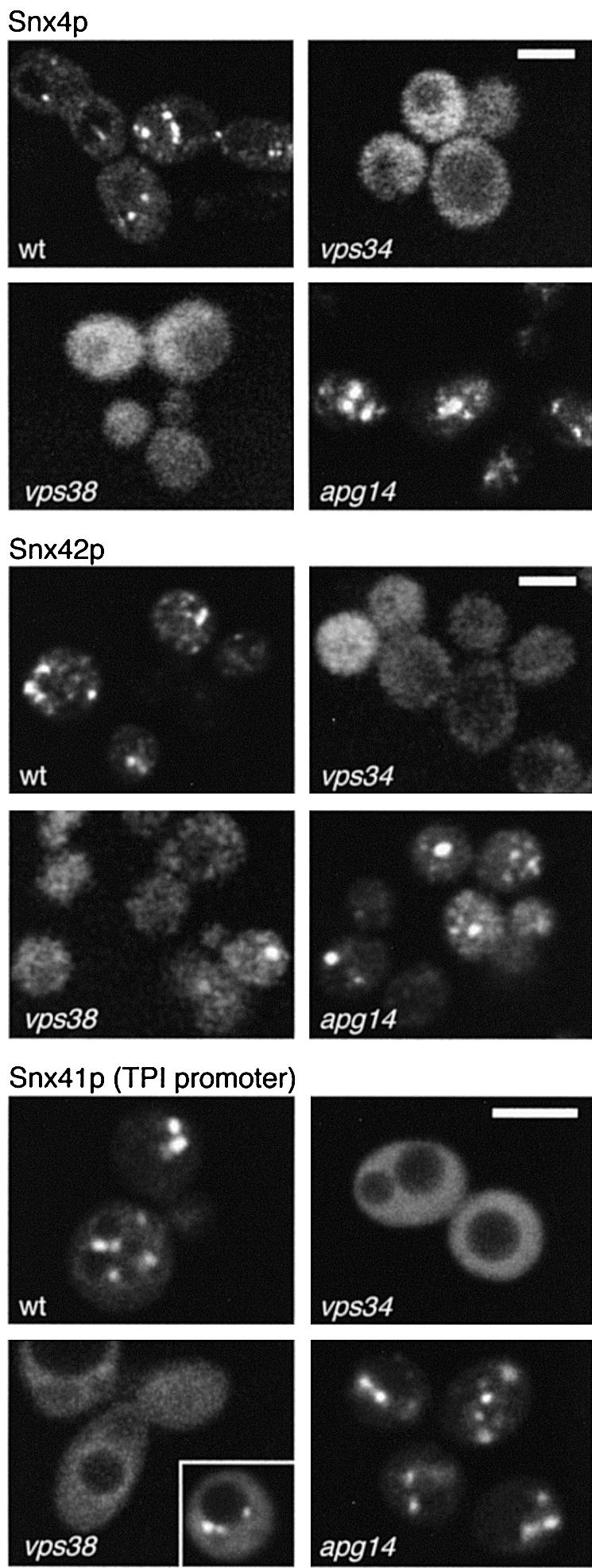
The endocytic pathway in yeast leads to the vacuole, but resident proteins of the late Golgi, and some endocytosed proteins such as the exocytic SNARE Snc1p, are retrieved specifically to the Golgi. Retrieval can occur from both a late pre-vacuolar compartment and early or 'post-Golgi' endosomes. We show that the endosomal SNARE Pep12p, and a mutant version that reaches the cell surface and is endocytosed, are retrieved from pre-vacuolar endosomes. As with Golgi proteins, this requires the sorting nexin Grd19p and components of the retromer coat, supporting the view that endosomal and Golgi residents both cycle continuously between the exocytic and endocytic pathways. In contrast, retrieval of Snc1p from post-Golgi endosomes requires the sorting nexin Snx4p, to which Snc1p can be cross-linked. Snx4p binds to Snx41p/ydr425w and to Snx42p/ydl113c, both of which are also required for efficient Snc1p sorting. Our findings suggest a general role for yeast sorting nexins in protein retrieval, rather than degradation, and indicate that different sorting nexins operate in different classes of endosomes.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
