

In vivo evidence for the prokaryotic model of extended codon-anticodon interaction in translation initiation
- PMID: 12554665
- PMCID: PMC140755
- DOI: 10.1093/emboj/cdg072
In vivo evidence for the prokaryotic model of extended codon-anticodon interaction in translation initiation
Abstract
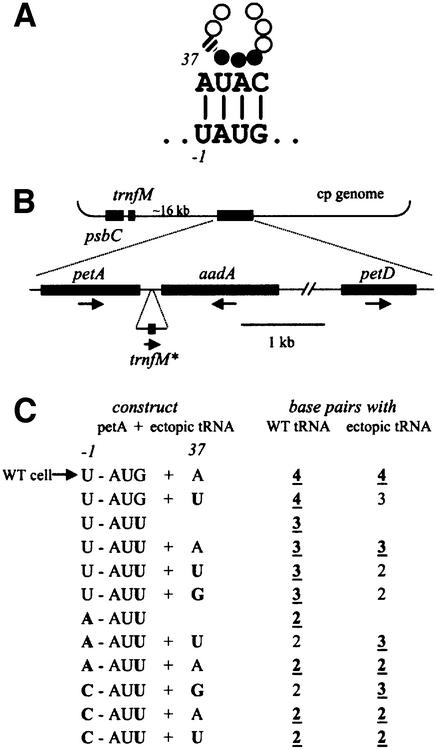
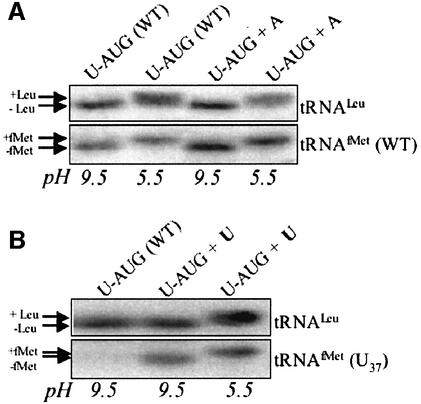
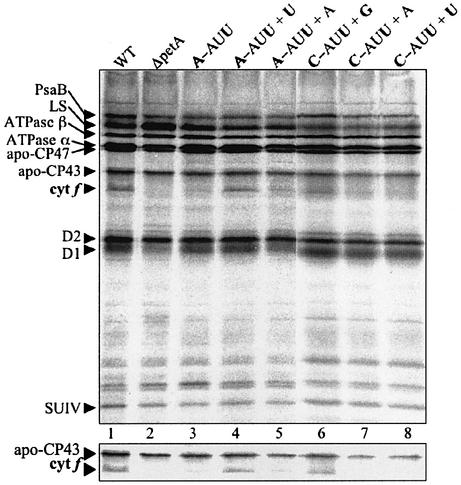
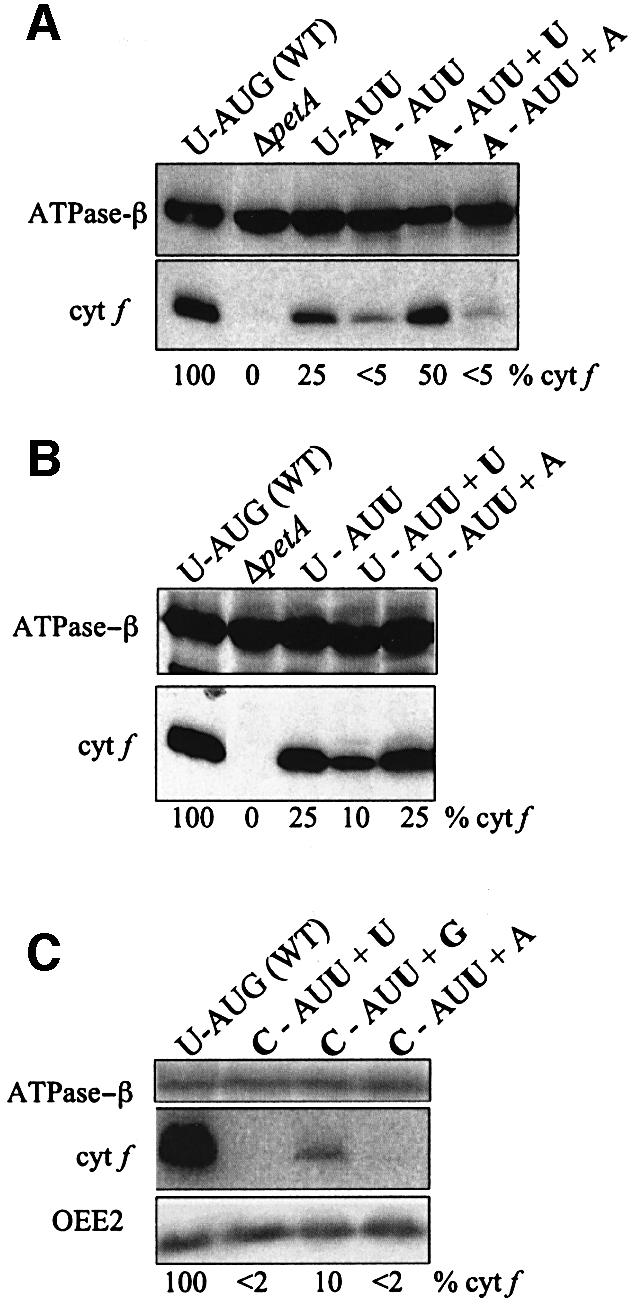
Initiation codon context is an important determinant of translation initiation rates in both prokaryotes and eukaryotes. Such sequences include the Shine- Dalgarno ribosome-binding site, as well as other motifs surrounding the initiation codon. One proposed interaction is between the base immediately preceding the initiation codon (-1 position) and the nucleotide 3' to the tRNAf(Met) anticodon, at position 37. Adenine is conserved at position 37, and a uridine at -1 has been shown in vitro to favor initiation. We have tested this model in vivo, by manipulating the chloroplast of the green alga Chlamydomonas reinhardtii, where the translational machinery is prokaryotic in nature. We show that translational defects imparted by mutations at the petA -1 position can be suppressed by compensatory mutations at position 37 of an ectopically expressed tRNA(fMet). The mutant tRNAs are fully aminoacylated and do not interfere with the translation of other proteins. Although this extended base pairing is not an absolute requirement for initiation, it may convey added specificity to transcripts carrying non-standard initiation codons, and/or preserve translational fidelity under certain stress conditions.
Figures
Similar articles
-
A role for initiation codon context in chloroplast translation.Plant Cell. 2001 Oct;13(10):2373-84. doi: 10.1105/tpc.010236. Plant Cell. 2001. PMID: 11595808 Free PMC article.
-
Compilation of mRNA sequences surrounding the AUG translation initiation codon in the green alga Chlamydomonas reinhardtii.Biosci Biotechnol Biochem. 1998 Dec;62(12):2457-9. doi: 10.1271/bbb.62.2457. Biosci Biotechnol Biochem. 1998. PMID: 9972275
-
Mechanisms governing codon usage bias and the implications for protein expression in the chloroplast of Chlamydomonas reinhardtii.Plant J. 2022 Nov;112(4):919-945. doi: 10.1111/tpj.15970. Epub 2022 Oct 19. Plant J. 2022. PMID: 36071273 Free PMC article. Review.
-
CITRIC: cold-inducible translational readthrough in the chloroplast of Chlamydomonas reinhardtii using a novel temperature-sensitive transfer RNA.Microb Cell Fact. 2018 Nov 24;17(1):186. doi: 10.1186/s12934-018-1033-5. Microb Cell Fact. 2018. PMID: 30474564 Free PMC article.
-
The ribosome's response to codon-anticodon mismatches.Biochimie. 2006 Aug;88(8):1001-11. doi: 10.1016/j.biochi.2006.04.013. Epub 2006 May 12. Biochimie. 2006. PMID: 16716484 Review.
Cited by
-
Overview of tRNA Modifications in Chloroplasts.Microorganisms. 2022 Jan 20;10(2):226. doi: 10.3390/microorganisms10020226. Microorganisms. 2022. PMID: 35208681 Free PMC article.
-
Evolution of monoblepharidalean fungi based on complete mitochondrial genome sequences.Nucleic Acids Res. 2003 Mar 15;31(6):1614-23. doi: 10.1093/nar/gkg264. Nucleic Acids Res. 2003. PMID: 12626702 Free PMC article.
-
Chloroplast Translation: Structural and Functional Organization, Operational Control, and Regulation.Plant Cell. 2018 Apr;30(4):745-770. doi: 10.1105/tpc.18.00016. Epub 2018 Apr 2. Plant Cell. 2018. PMID: 29610211 Free PMC article. Review.
-
Development of novel types of plastid transformation vectors and evaluation of factors controlling expression.Transgenic Res. 2005 Dec;14(6):969-82. doi: 10.1007/s11248-005-2542-7. Transgenic Res. 2005. PMID: 16315098
-
Regulatory sequences of orthologous petD chloroplast mRNAs are highly specific among Chlamydomonas species.Plant Mol Biol. 2006 Feb;60(3):405-22. doi: 10.1007/s11103-005-4477-z. Plant Mol Biol. 2006. PMID: 16514563
References
-
- Culbertson M., Leeds,P., Sandbaken,M. and Wilson,P. (1990) Frameshift suppression. In Hill,W., Dahlberg,A., Garrett,P., Moore,P., Schlessinger,D. and Warner,J. (eds), The Ribosome: Structure, Function and Evolution. American Society for Microbiology, Washington, DC, pp. 559–570.
-
- Delamarche C., Guerdoux-Jamet,P., Gras,R. and Nicolas,J. (1999) A symbolic–numeric approach to find patterns in genomes. Application to the translation initiation sites of E.coli. Biochimie, 81, 1065–1072. - PubMed
-
- Drager R.G., Girard-Bascou,J., Choquet,Y., Kindle,K.L. and Stern,D.B. (1998) In vivo evidence for 5′→3′ exoribonuclease degradation of an unstable chloroplast mRNA. Plant J., 13, 85–96. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
