

A PIT-1 homeodomain mutant blocks the intranuclear recruitment of the CCAAT/enhancer binding protein alpha required for prolactin gene transcription
- PMID: 12554749
- PMCID: PMC2900764
- DOI: 10.1210/me.2001-0222
A PIT-1 homeodomain mutant blocks the intranuclear recruitment of the CCAAT/enhancer binding protein alpha required for prolactin gene transcription
Abstract
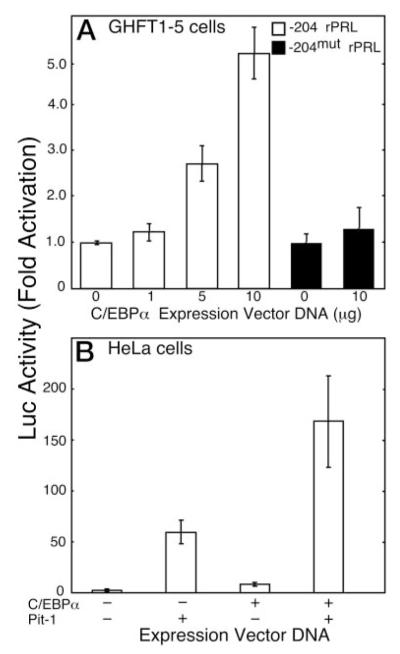
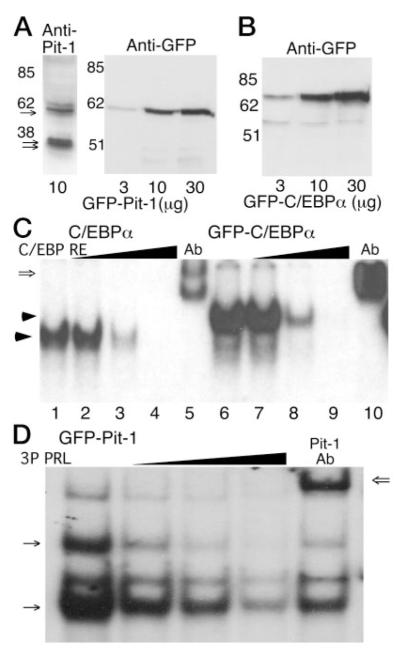
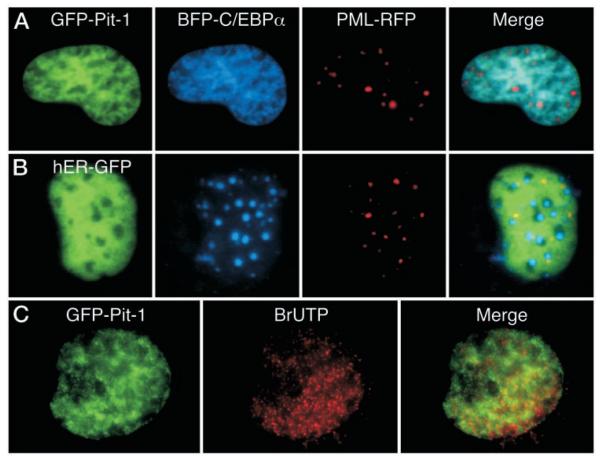
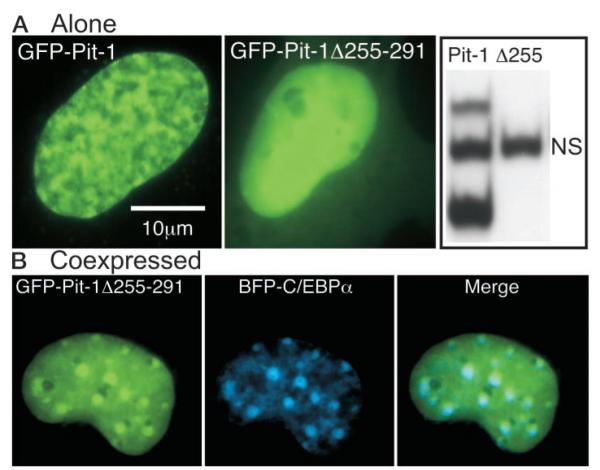
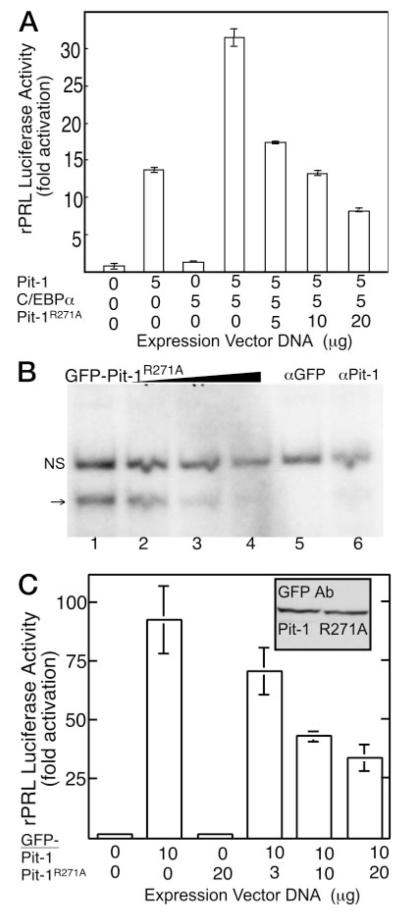
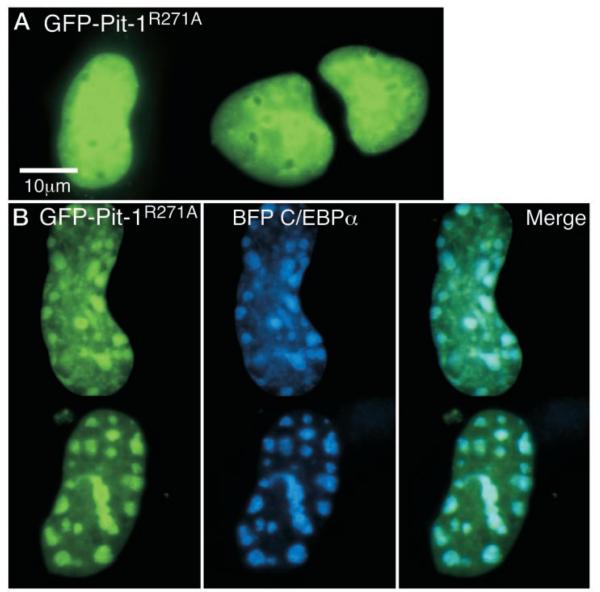
The pituitary-specific homeodomain protein Pit-1 cooperates with other transcription factors, including CCAAT/enhancer binding protein alpha (C/EBPalpha), in the regulation of pituitary lactotrope gene transcription. Here, we correlate cooperative activation of prolactin (PRL) gene transcription by Pit-1 and C/EBPalpha with changes in the subnuclear localization of these factors in living pituitary cells. Transiently expressed C/EBPalpha induced PRL gene transcription in pituitary GHFT1-5 cells, whereas the coexpression of Pit-1 and C/EBPalpha in HeLa cells demonstrated their cooperativity at the PRL promoter. Individually expressed Pit-1 or C/EBPalpha, fused to color variants of fluorescent proteins, occupied different subnuclear compartments in living pituitary cells. When coexpressed, Pit-1 recruited C/EBPalpha from regions of transcriptionally quiescent centromeric heterochromatin to the nuclear regions occupied by Pit-1. The homeodomain region of Pit-1 was necessary for the recruitment of C/EBPalpha. A point mutation in the Pit-1 homeodomain associated with the syndrome of combined pituitary hormone deficiency in humans also failed to recruit C/EBPalpha. This Pit-1 mutant functioned as a dominant inhibitor of PRL gene transcription and, instead of recruiting C/EBPalpha, was itself recruited by C/EBPalpha to centromeric heterochromatin. Together our results suggest that the intranuclear positioning of these factors determines whether they activate or silence PRL promoter activity.
Figures

Similar articles
-
Imaging the localized protein interactions between Pit-1 and the CCAAT/enhancer binding protein alpha in the living pituitary cell nucleus.Mol Endocrinol. 2003 Mar;17(3):333-45. doi: 10.1210/me.2002-0136. Epub 2002 Dec 12. Mol Endocrinol. 2003. PMID: 12554785 Free PMC article.
-
Dynamic interactions between Pit-1 and C/EBPalpha in the pituitary cell nucleus.Mol Cell Biol. 2006 Nov;26(21):8087-98. doi: 10.1128/MCB.02410-05. Epub 2006 Aug 14. Mol Cell Biol. 2006. PMID: 16908544 Free PMC article.
-
CCAAT/enhancer binding protein alpha uses distinct domains to prolong pituitary cells in the growth 1 and DNA synthesis phases of the cell cycle.BMC Cell Biol. 2002 Mar 21;3:6. doi: 10.1186/1471-2121-3-6. BMC Cell Biol. 2002. PMID: 11914124 Free PMC article.
-
Role of Pit-1 in the gene expression of growth hormone, prolactin, and thyrotropin.Endocrinol Metab Clin North Am. 1996 Sep;25(3):523-40. doi: 10.1016/s0889-8529(05)70339-x. Endocrinol Metab Clin North Am. 1996. PMID: 8879985 Review.
-
POU domain factors in neural development.Adv Exp Med Biol. 1998;449:39-53. doi: 10.1007/978-1-4615-4871-3_4. Adv Exp Med Biol. 1998. PMID: 10026784 Review.
Cited by
-
Computer-assisted image analysis protocol that quantitatively measures subnuclear protein organization in cell populations.Biotechniques. 2004 Feb;36(2):240-7. doi: 10.2144/04362BI01. Biotechniques. 2004. PMID: 14989088 Free PMC article.
-
Regulation of LH/FSH expression by secretoglobin 3A2 in the mouse pituitary gland.Cell Tissue Res. 2014 Apr;356(1):253-60. doi: 10.1007/s00441-014-1794-z. Epub 2014 Feb 11. Cell Tissue Res. 2014. PMID: 24514953 Free PMC article.
-
Imaging the localized protein interactions between Pit-1 and the CCAAT/enhancer binding protein alpha in the living pituitary cell nucleus.Mol Endocrinol. 2003 Mar;17(3):333-45. doi: 10.1210/me.2002-0136. Epub 2002 Dec 12. Mol Endocrinol. 2003. PMID: 12554785 Free PMC article.
-
The many faces of C/EBPδ and their relevance for inflammation and cancer.Int J Biol Sci. 2013 Sep 20;9(9):917-33. doi: 10.7150/ijbs.7224. eCollection 2013. Int J Biol Sci. 2013. PMID: 24155666 Free PMC article. Review.
-
Functional sequestration of transcription factor activity by repetitive DNA.J Biol Chem. 2007 Jul 20;282(29):20868-76. doi: 10.1074/jbc.M702547200. Epub 2007 May 25. J Biol Chem. 2007. PMID: 17526489 Free PMC article.
References
-
- FJ Iborra, DA Jackson, PR Cook. The path of transcripts from extra-nucleolar synthetic sites to nuclear pores: transcripts in transit are concentrated in discrete structures containing SR proteins. J Cell Sci. 1996;111:2269–2282. - PubMed
-
- TA Nardozza, MM Quigley, RH Getzenberg. Association of transcription factors with the nuclear matrix. J Cell Biochem. 1996;61:467–477. - PubMed
-
- AI Lamond, WC Earnshaw. Structure and function in the nucleus. Science. 1998;280:547–553. - PubMed
-
- A Pombo, P Cuello, W Schul, JB Yoon, RG Roeder, PR Cook, S Murphy. Regional and temporal specialization in the nucleus: a transcriptionally-active nuclear domain rich in PTF, Oct1 and PIKA antigens associates with specific chromosomes early in the cell cycle. EMBO J. 1998;17:1768–1778. - PMC - PubMed
-
- T Misteli. Protein dynamics: implications for nuclear architecture and gene expression. Science. 2001;291:843–847. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
