

Nuclear translation: what is the evidence?
- PMID: 12554869
- PMCID: PMC1370363
- DOI: 10.1261/rna.2121703
Nuclear translation: what is the evidence?
Abstract
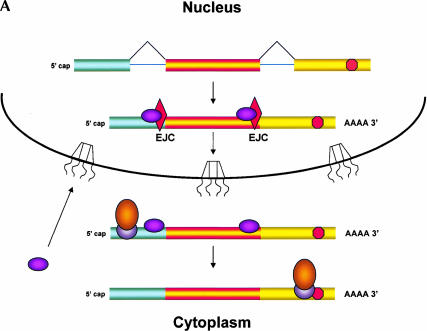
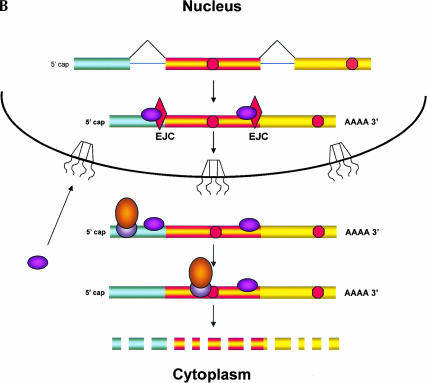
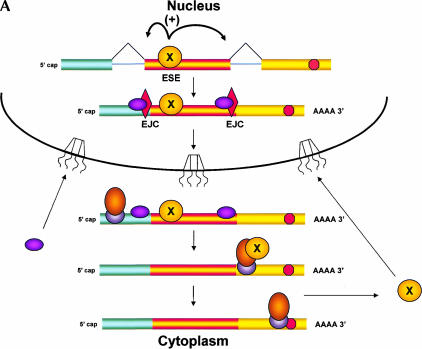
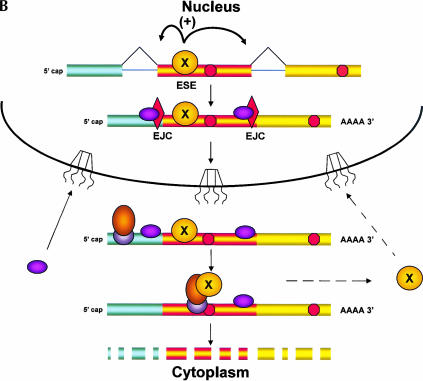
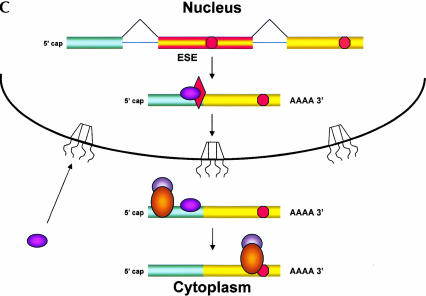
Recently, several reports have been published in support of the idea that protein synthesis occurs in both the nucleus and the cytoplasm. This proposal has generated a great deal of excitement because, if true, it would mean that our thinking about the compartmentalization of cell functions would have to be re-evaluated. The significance and broad implications of this phenomenon require that the experimental evidence used to support it be carefully evaluated. Here, we critique the published evidence in support of, or in opposition to, the question of whether translation occurs in the nucleus. Arguments in support of nuclear translation focus on three issues: (1) the presence of translation factors and ribosomal components in the nucleus, and their recruitment to sites of transcription; (2) amino acid incorporation in isolated nuclei and in nuclei under conditions that should not permit protein import; and (3) the fact that nuclear translation would account for observations that are otherwise difficult to explain. Arguments against nuclear translation emphasize the absence (or low abundance) from nuclei of many translation factors; the likely inactivity of nascent ribosomes; and the loss of translation activity as nuclei are purified from contaminating cytoplasm. In our opinion, all of the experiments on nuclear translation published to date lack critical controls and, therefore, are not compelling; also, traditional mechanisms can explain the observations for which nuclear translation has been invoked. Thus, while we cannot rule out nuclear translation, in the absence of better supporting data we are reluctant to believe it occurs.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources