

Identification of the Hfq-binding site on DsrA RNA: Hfq binds without altering DsrA secondary structure
- PMID: 12554874
- PMCID: PMC1370368
- DOI: 10.1261/rna.2570803
Identification of the Hfq-binding site on DsrA RNA: Hfq binds without altering DsrA secondary structure
Abstract
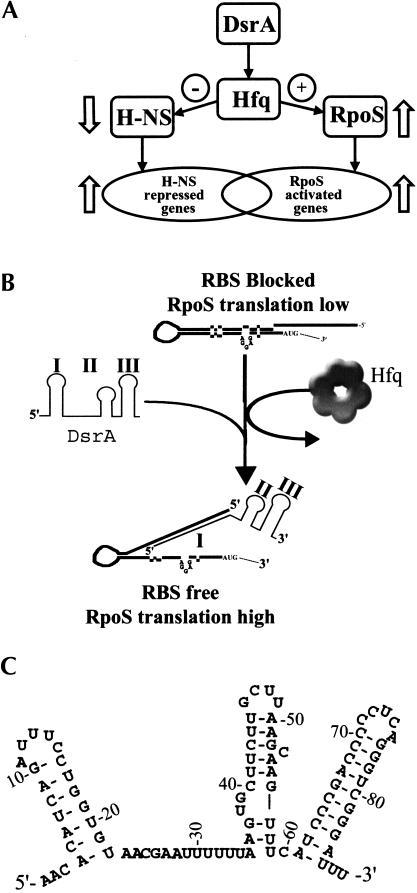
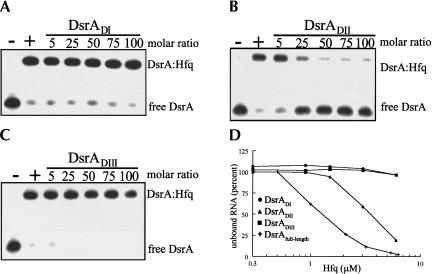
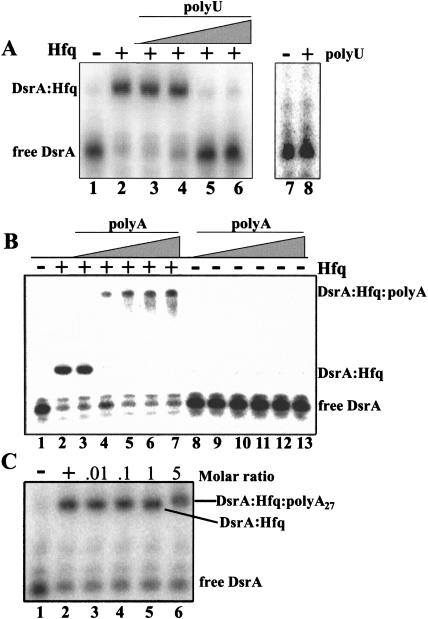
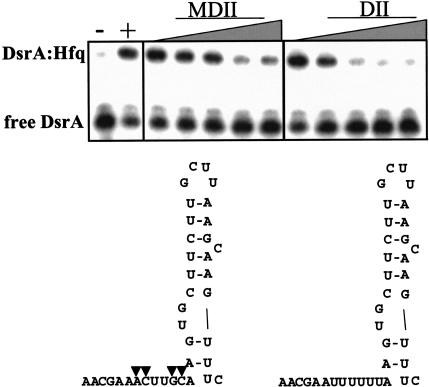
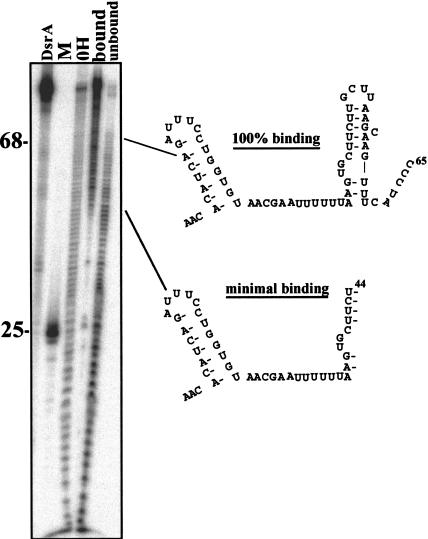
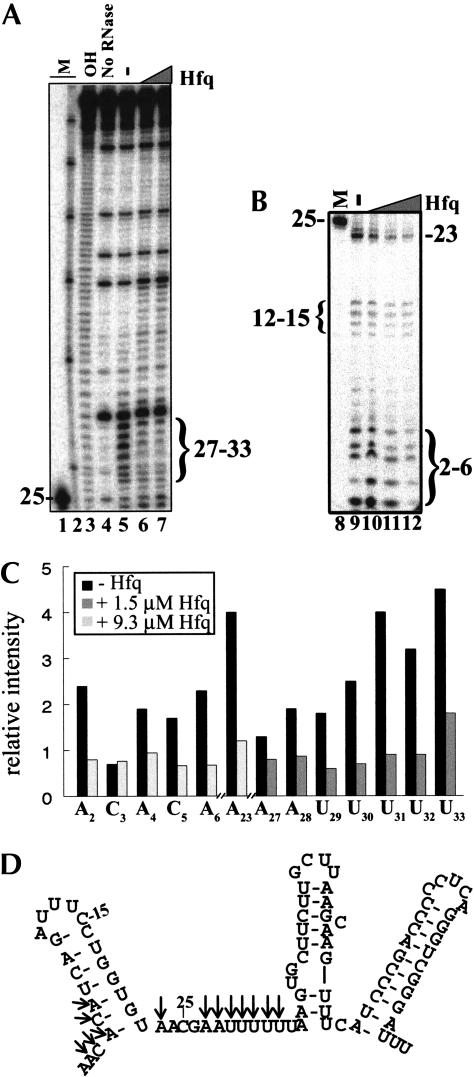
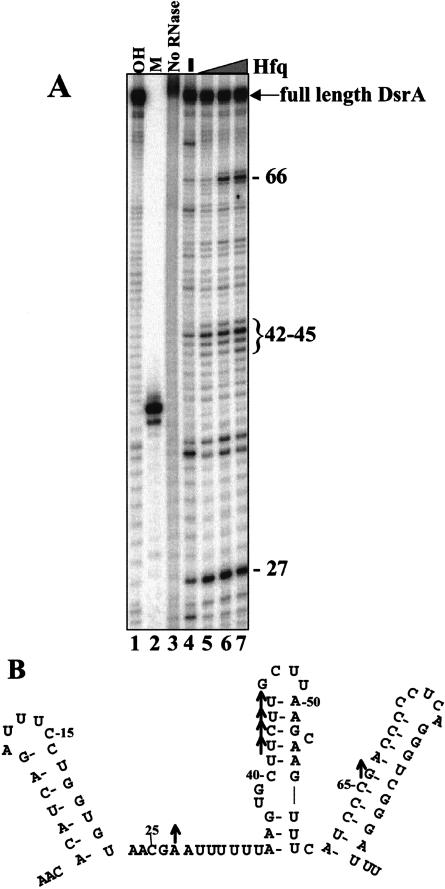
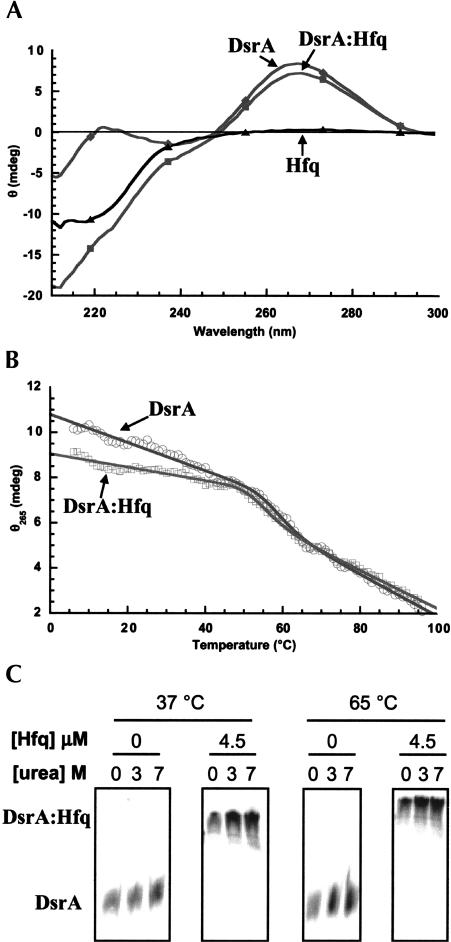
DsrA RNA regulates the translation of two global regulatory proteins in Escherichia coli. DsrA activates the translation of RpoS while repressing the translation of H-NS. The RNA-binding protein Hfq is necessary for DsrA to function in vivo. Although Hfq binds to DsrA in vitro, the role of Hfq in DsrA-mediated regulation is not known. One hypothesis was that Hfq acts as an RNA chaperone by unfolding DsrA, thereby facilitating interactions with target RNAs. To test this hypothesis, we have examined the structure of DsrA bound to Hfq in vitro. Comparison of free DsrA to DsrA bound to Hfq by RNase footprinting, circular dichroism, and thermal melt profiles shows that Hfq does not alter DsrA secondary structures, but might affect its tertiary conformation. We identify the site on DsrA where Hfq binds, which is a structural element in the middle of DsrA. In addition, we show that although long poly(U) RNAs compete with DsrA for binding to Hfq, a short poly(U) stretch present in DsrA is not necessary for Hfq binding. Finally, unlike other RNAs, DsrA binding to Hfq is not competed with by poly(A) RNA. In fact, DsrA:poly(A):Hfq may form a stable ternary complex, raising the possibility that Hfq has multiple RNA-binding sites.
Figures
References
-
- Arluison, V., Derreumaux, P., Allemand, F., Folichon, M., Hajnsdorf, E., and Regnier, P. 2002. Structural modelling of the Sm-like Protein Hfq from Escherichia coli. J. Mol. Biol. 320: 705–712. - PubMed
-
- Bloomfield, V.A., Crothers, D.M., and Tinoco, J.I. 2000. Nucleic acids: Structures, properties and function. University Science Books, Sausalito, CA.
-
- Collins, B.M., Harrop, S.J., Kornfeld, G.D., Dawes, I.W., Curmi, P.M., and Mabbutt, B.C. 2001. Crystal structure of a heptameric Sm-like protein complex from archaea: Implications for the structure and evolution of snRNPs. J. Mol. Biol. 309: 915–923. - PubMed
-
- de Haseth, P.L. and Uhlenbeck, O.C. 1980a. Interaction of Escherichia coli host factor protein with oligoriboadenylates. Biochemistry 19: 6138–6146. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous