

batman Interacts with polycomb and trithorax group genes and encodes a BTB/POZ protein that is included in a complex containing GAGA factor
- PMID: 12556479
- PMCID: PMC141128
- DOI: 10.1128/MCB.23.4.1181-1195.2003
batman Interacts with polycomb and trithorax group genes and encodes a BTB/POZ protein that is included in a complex containing GAGA factor
Abstract
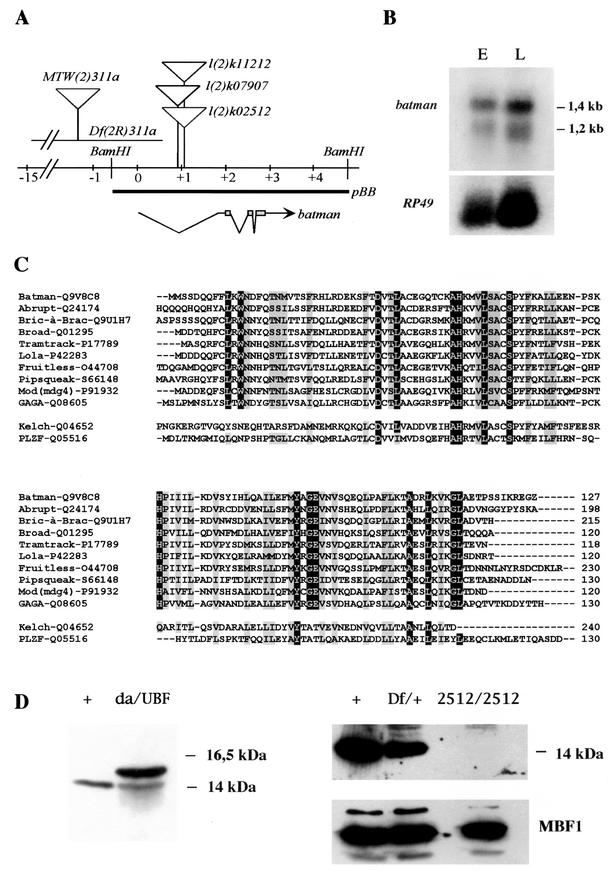
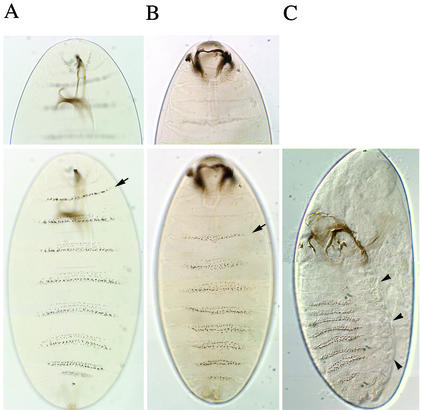
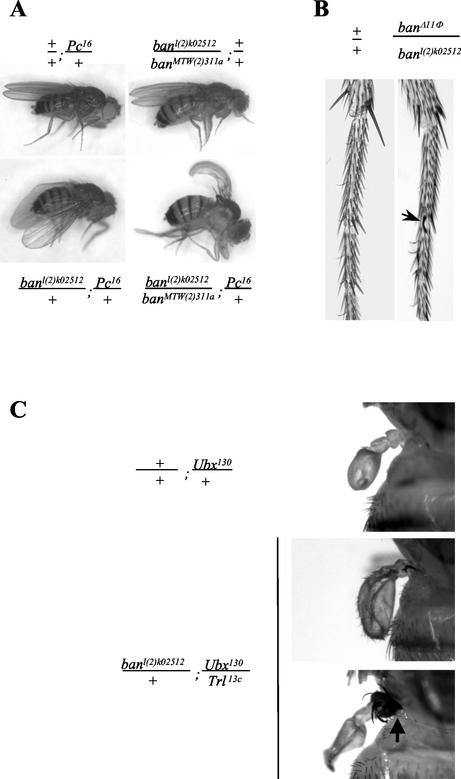
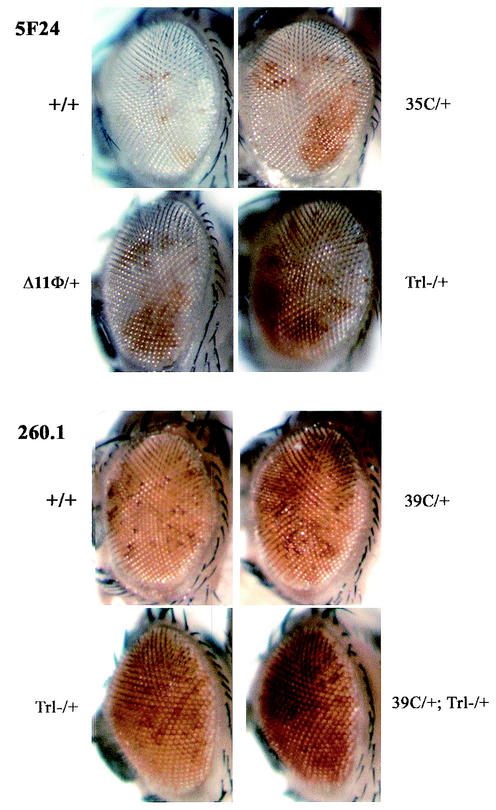
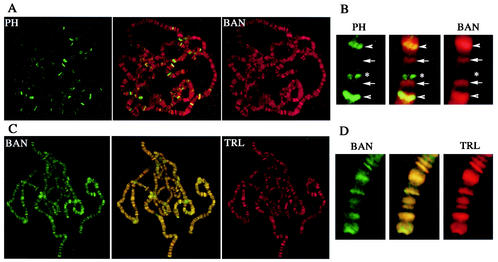
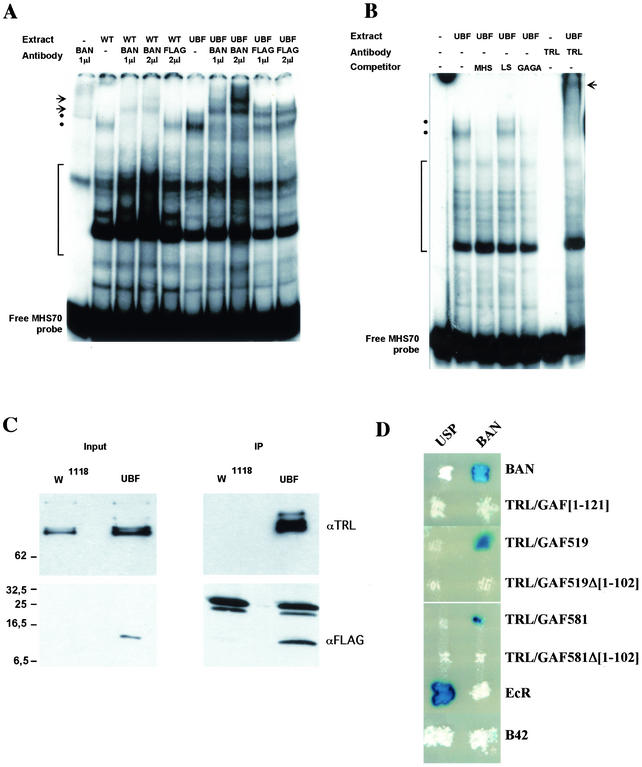
Polycomb and trithorax group genes maintain the appropriate repressed or activated state of homeotic gene expression throughout Drosophila melanogaster development. We have previously identified the batman gene as a Polycomb group candidate since its function is necessary for the repression of Sex combs reduced. However, our present genetic analysis indicates functions of batman in both activation and repression of homeotic genes. The 127-amino-acid Batman protein is almost reduced to a BTB/POZ domain, an evolutionary conserved protein-protein interaction domain found in a large protein family. We show that this domain is involved in the interaction between Batman and the DNA binding GAGA factor encoded by the Trithorax-like gene. The GAGA factor and Batman codistribute on polytene chromosomes, coimmunoprecipitate from nuclear embryonic and larval extracts, and interact in the yeast two-hybrid assay. Batman, together with the GAGA factor, binds to MHS-70, a 70-bp fragment of the bithoraxoid Polycomb response element. This binding, like that of the GAGA factor, requires the presence of d(GA)n sequences. Together, our results suggest that batman belongs to a subset of the Polycomb/trithorax group of genes that includes Trithorax-like, whose products are involved in both activation and repression of homeotic genes.
Figures
Similar articles
-
The Drosophila pho-like gene encodes a YY1-related DNA binding protein that is redundant with pleiohomeotic in homeotic gene silencing.Development. 2003 Jan;130(2):285-94. doi: 10.1242/dev.00204. Development. 2003. PMID: 12466196
-
Regulation of Polycomb group complexes by the sequence-specific DNA binding proteins Zeste and GAGA.Genes Dev. 2003 Nov 15;17(22):2741-6. doi: 10.1101/gad.1143303. Genes Dev. 2003. PMID: 14630938 Free PMC article.
-
The Drosophila Polycomb group gene Sex combs extra encodes the ortholog of mammalian Ring1 proteins.Mech Dev. 2004 May;121(5):449-62. doi: 10.1016/j.mod.2004.03.019. Mech Dev. 2004. PMID: 15147763
-
The Polycomb and trithorax group proteins of Drosophila: trans-regulators of homeotic gene function.Annu Rev Genet. 1995;29:289-303. doi: 10.1146/annurev.ge.29.120195.001445. Annu Rev Genet. 1995. PMID: 8825476 Review.
-
Epigenetic inheritance of chromatin states mediated by Polycomb and trithorax group proteins in Drosophila.Prog Mol Subcell Biol. 2005;38:31-63. doi: 10.1007/3-540-27310-7_2. Prog Mol Subcell Biol. 2005. PMID: 15881890 Review.
Cited by
-
The Arabidopsis GAGA-Binding Factor BASIC PENTACYSTEINE6 Recruits the POLYCOMB-REPRESSIVE COMPLEX1 Component LIKE HETEROCHROMATIN PROTEIN1 to GAGA DNA Motifs.Plant Physiol. 2015 Jul;168(3):1013-24. doi: 10.1104/pp.15.00409. Epub 2015 May 29. Plant Physiol. 2015. PMID: 26025051 Free PMC article.
-
lolal Is an Evolutionarily New Epigenetic Regulator of dpp Transcription during Dorsal-Ventral Axis Formation.Mol Biol Evol. 2016 Oct;33(10):2621-32. doi: 10.1093/molbev/msw132. Epub 2016 Jul 8. Mol Biol Evol. 2016. PMID: 27401231 Free PMC article.
-
Nuclear import of transcription factor BR-C is mediated by its interaction with RACK1.PLoS One. 2014 Oct 3;9(10):e109111. doi: 10.1371/journal.pone.0109111. eCollection 2014. PLoS One. 2014. PMID: 25280016 Free PMC article.
-
Ribbon modulates apical membrane during tube elongation through Crumbs and Moesin.Dev Biol. 2008 Aug 1;320(1):278-88. doi: 10.1016/j.ydbio.2008.05.541. Epub 2008 May 22. Dev Biol. 2008. PMID: 18585700 Free PMC article.
-
Phenotypic plasticity in Drosophila pigmentation caused by temperature sensitivity of a chromatin regulator network.PLoS Genet. 2007 Feb 16;3(2):e30. doi: 10.1371/journal.pgen.0030030. PLoS Genet. 2007. PMID: 17305433 Free PMC article.
References
-
- Albagli, O., P. Dhordain, C. Deweindt, G. Lecocq, and D. Leprince. 1995. The BTB/POZ domain: a new protein-protein interaction motif common to DNA- and actin-binding proteins. Cell Growth Differ. 6:1193-1198. - PubMed
-
- Aravind, L., and E. V. Koonin. 1999. Fold prediction and evolutionary analysis of the POZ domain: structural and evolutionary relationship with the potassium channel tetramerization domain. J. Mol. Biol. 285:1353-1361. - PubMed
-
- Bardwell, V. J., and R. Treisman. 1994. The POZ domain: a conserved protein-protein interaction motif. Genes Dev. 8:1664-1677. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases