

Formation of a tissue-specific histone acetylation pattern by the hematopoietic transcription factor GATA-1
- PMID: 12556492
- PMCID: PMC141148
- DOI: 10.1128/MCB.23.4.1334-1340.2003
Formation of a tissue-specific histone acetylation pattern by the hematopoietic transcription factor GATA-1
Abstract
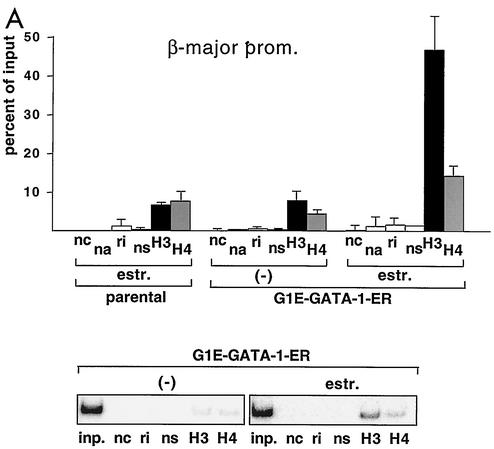
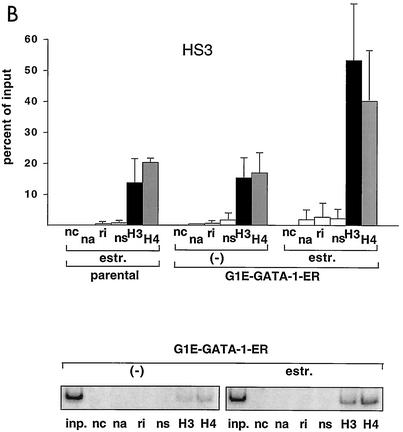
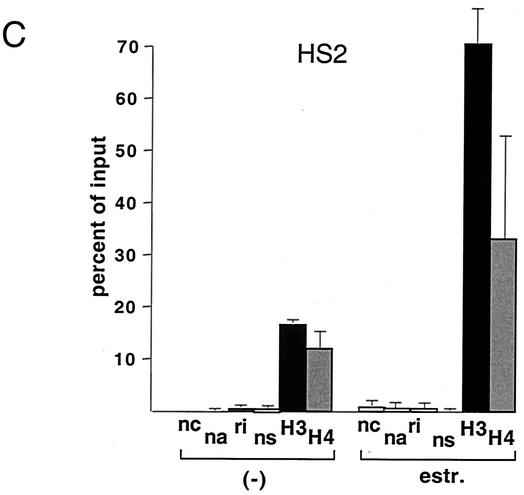
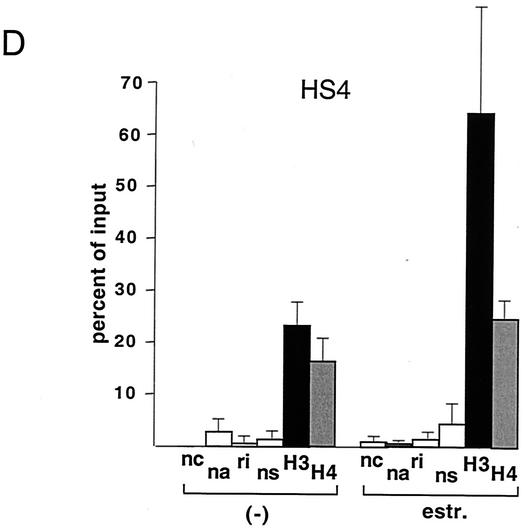
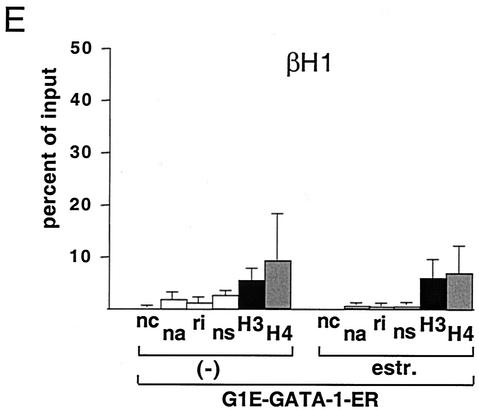
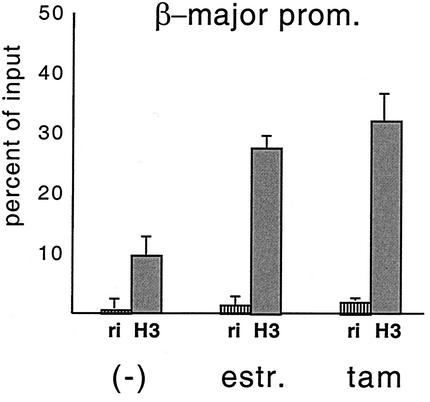
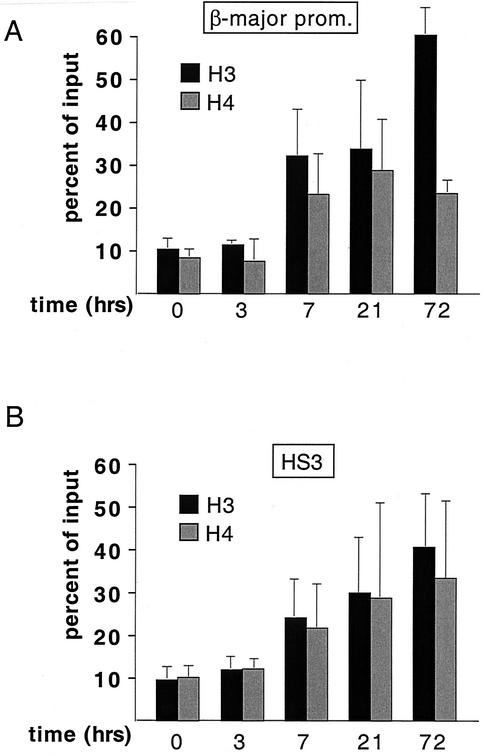
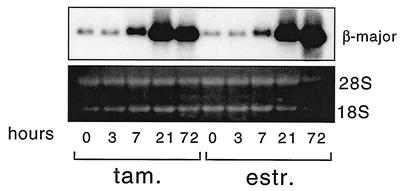
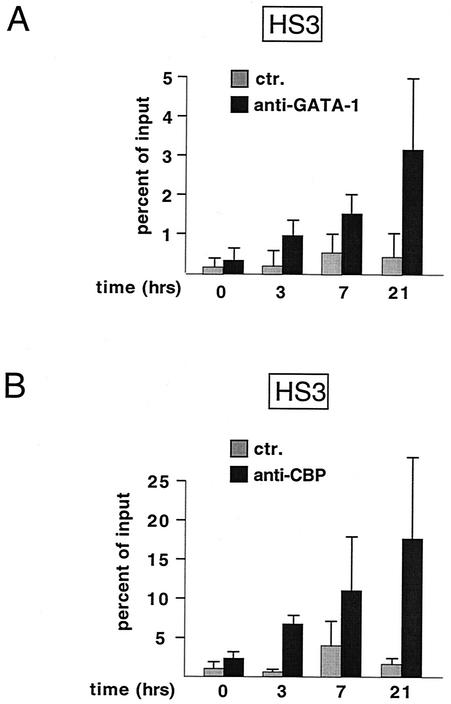
One function of lineage-restricted transcription factors may be to control the formation of tissue-specific chromatin domains. In erythroid cells, the beta-globin gene cluster undergoes developmentally regulated hyperacetylation of histones at the active globin genes and the locus control region (LCR). However, it is unknown which transcription factor(s) governs the establishment of this erythroid-specific chromatin domain. We measured histone acetylation at the beta-globin locus in the erythroid cell line G1E, which is deficient for the essential hematopoietic transcription factor GATA-1. Restoration of GATA-1 activity in G1E cells led to a substantial increase in acetylation of histones H3 and H4 at the beta-globin promoter and the LCR. Time course experiments showed that histone acetylation occurred rapidly after GATA-1 activation and coincided with globin gene expression, indicating that the effects of GATA-1 are direct. Moreover, increases in histone acetylation correlated with occupancy of GATA-1 and the acetyltransferase CBP at the locus in vivo. Together, these results suggest that GATA-1 and its cofactor CBP are essential for the formation of an erythroid-specific acetylation pattern that is permissive for high levels of gene expression.
Figures
Similar articles
-
Hematopoietic-specific activators establish an overlapping pattern of histone acetylation and methylation within a mammalian chromatin domain.Proc Natl Acad Sci U S A. 2002 Oct 29;99(22):14309-14. doi: 10.1073/pnas.212389499. Epub 2002 Oct 11. Proc Natl Acad Sci U S A. 2002. PMID: 12379744 Free PMC article.
-
Expression of GATA-1 in a non-hematopoietic cell line induces beta-globin locus control region chromatin structure remodeling and an erythroid pattern of gene expression.J Mol Biol. 2007 Feb 23;366(3):737-44. doi: 10.1016/j.jmb.2006.11.094. Epub 2006 Dec 6. J Mol Biol. 2007. PMID: 17196618 Free PMC article.
-
A WW domain-binding motif within the activation domain of the hematopoietic transcription factor NF-E2 is essential for establishment of a tissue-specific histone modification pattern.J Biol Chem. 2004 Feb 27;279(9):7456-61. doi: 10.1074/jbc.M309750200. Epub 2003 Nov 2. J Biol Chem. 2004. PMID: 14597626
-
ChIPs of the beta-globin locus: unraveling gene regulation within an active domain.Curr Opin Genet Dev. 2002 Apr;12(2):170-7. doi: 10.1016/s0959-437x(02)00283-6. Curr Opin Genet Dev. 2002. PMID: 11893490 Review.
-
Transcription factor GATA-1 and erythroid development.Proc Soc Exp Biol Med. 1993 Feb;202(2):115-21. doi: 10.3181/00379727-202-43519a. Proc Soc Exp Biol Med. 1993. PMID: 8424101 Review.
Cited by
-
Differential sensitivities of transcription factor target genes underlie cell type-specific gene expression profiles.Proc Natl Acad Sci U S A. 2006 Oct 24;103(43):15939-44. doi: 10.1073/pnas.0604041103. Epub 2006 Oct 16. Proc Natl Acad Sci U S A. 2006. PMID: 17043224 Free PMC article.
-
A global role for EKLF in definitive and primitive erythropoiesis.Blood. 2006 Apr 15;107(8):3359-70. doi: 10.1182/blood-2005-07-2888. Epub 2005 Dec 27. Blood. 2006. PMID: 16380451 Free PMC article.
-
Context-dependent regulation of GATA-1 by friend of GATA-1.Proc Natl Acad Sci U S A. 2004 Jan 13;101(2):476-81. doi: 10.1073/pnas.0306315101. Epub 2003 Dec 26. Proc Natl Acad Sci U S A. 2004. PMID: 14695898 Free PMC article.
-
Bromodomain protein Brd3 associates with acetylated GATA1 to promote its chromatin occupancy at erythroid target genes.Proc Natl Acad Sci U S A. 2011 May 31;108(22):E159-68. doi: 10.1073/pnas.1102140108. Epub 2011 May 2. Proc Natl Acad Sci U S A. 2011. PMID: 21536911 Free PMC article.
-
GATA4 represses an ileal program of gene expression in the proximal small intestine by inhibiting the acetylation of histone H3, lysine 27.Biochim Biophys Acta. 2014 Nov;1839(11):1273-82. doi: 10.1016/j.bbagrm.2014.05.018. Epub 2014 May 27. Biochim Biophys Acta. 2014. PMID: 24878542 Free PMC article.
References
-
- Bender, M. A., M. Bulger, J. Close, and M. Groudine. 2000. Beta-globin gene switching and DNase I sensitivity of the endogenous beta-globin locus in mice do not require the locus control region. Mol. Cell 5:387-393. - PubMed
-
- Blobel, G. A. 2000. CBP/p300: molecular integrators of hematopoietic transcription. Blood 95:745-755. - PubMed
-
- Bulger, M., T. Sawado, D. Schubeler, and M. Groudine. 2002. ChIPs of the beta-globin locus: unraveling gene regulation within an active domain. Curr. Opin. Genet. Dev. 12:170-177. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources