

Mathematical models of hierarchically structured cell populations under equilibrium with application to the epidermis
- PMID: 12558657
- PMCID: PMC6734918
- DOI: 10.1046/j.1365-2184.2003.00257.x
Mathematical models of hierarchically structured cell populations under equilibrium with application to the epidermis
Abstract
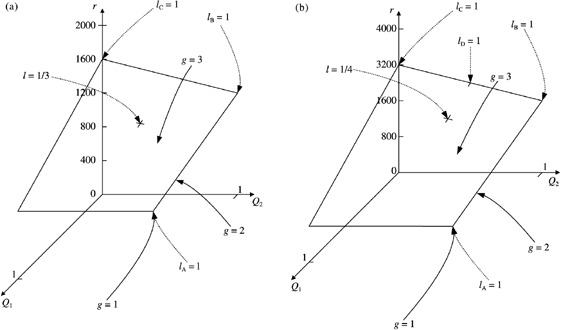
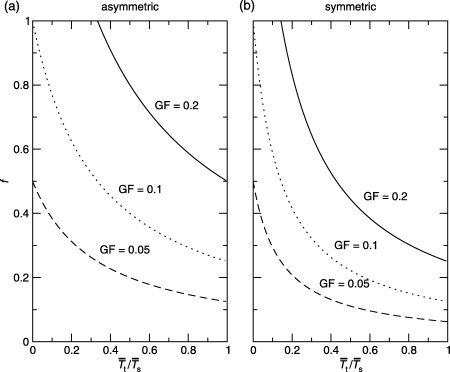
There are three categories of keratinocytes in the germinative compartment of the epidermis - stem, transit-amplifying and post-mitotic. Their population structure is hierarchical. This means that stem cells differentiate into transit-amplifying cells which, after a few rounds of division, become post-mitotic cells. The cell processes of birth, differentiation, death and migration affect the composition and proliferation rate of the germinative compartment. These phenomena are quantified by various cell kinetic parameters. In this paper we derive equations that relate these parameters for different models of hierarchically structured cell populations in equilibrium. We include in the models asymmetric and symmetric division, variations in cell-cycle times, apoptosis and variation in the number of transit generations. We conclude that variation in cell-cycle times need only be considered if apoptosis is not negligible. If it is negligible, then only average cell-cycle times are needed. Unfortunately, it is impossible to predict the importance of apoptosis from the available experimental data. However, the strength of its effect is determined by the other parameters, especially the fraction of cycling stem cells. We show that variation in the number of transit generations can have a potentially large effect on cell birth rate. We also show that cell birth rate does not directly depend on the mean transit-amplifying cell-cycle time, only on the mean stem cell-cycle time. We argue that 'homogeneous cell population' equations should not be used to study hierarchical cell populations as has been done in the past. Finally we argue that stem cell parameters and transit-amplifying cell parameters should not be lumped together.
Figures
Similar articles
-
c-Myc promotes differentiation of human epidermal stem cells.Genes Dev. 1997 Nov 1;11(21):2869-82. doi: 10.1101/gad.11.21.2869. Genes Dev. 1997. PMID: 9353256 Free PMC article.
-
Epidermal stem cells: markers, patterning and the control of stem cell fate.Philos Trans R Soc Lond B Biol Sci. 1998 Jun 29;353(1370):831-7. doi: 10.1098/rstb.1998.0247. Philos Trans R Soc Lond B Biol Sci. 1998. PMID: 9684280 Free PMC article. Review.
-
The growth fraction of normal human epidermis.Dermatology. 1997;194(4):313-7. doi: 10.1159/000246122. Dermatology. 1997. PMID: 9252749 Review.
-
Simulating psoriasis by altering transit amplifying cells.Bioinformatics. 2007 Jun 1;23(11):1309-12. doi: 10.1093/bioinformatics/btm042. Epub 2007 Feb 18. Bioinformatics. 2007. PMID: 17308343
-
Separation of human epidermal stem cells from transit amplifying cells on the basis of differences in integrin function and expression.Cell. 1993 May 21;73(4):713-24. doi: 10.1016/0092-8674(93)90251-k. Cell. 1993. PMID: 8500165
Cited by
-
An integrated systems biology approach to understanding the rules of keratinocyte colony formation.J R Soc Interface. 2007 Dec 22;4(17):1077-92. doi: 10.1098/rsif.2007.0227. J R Soc Interface. 2007. PMID: 17374590 Free PMC article.
-
Epidermal kinetic alterations required to generate the psoriatic phenotype: a reappraisal.Cell Prolif. 2010 Jun;43(3):321-5. doi: 10.1111/j.1365-2184.2010.00672.x. Cell Prolif. 2010. PMID: 20546247 Free PMC article.
-
Robustness of differentiation cascades with symmetric stem cell division.J R Soc Interface. 2014 Apr 9;11(95):20140264. doi: 10.1098/rsif.2014.0264. Print 2014 Jun 6. J R Soc Interface. 2014. PMID: 24718457 Free PMC article.
-
On Discretely Structured Growth Models and Their Moments.Bull Math Biol. 2025 May 12;87(6):71. doi: 10.1007/s11538-025-01446-w. Bull Math Biol. 2025. PMID: 40353893 Free PMC article.
-
Cardiomyogenesis in the developing heart is regulated by c-kit-positive cardiac stem cells.Circ Res. 2012 Mar 2;110(5):701-15. doi: 10.1161/CIRCRESAHA.111.259507. Epub 2012 Jan 24. Circ Res. 2012. Retraction in: Circ Res. 2019 Feb 15;124(4):e28. doi: 10.1161/RES.0000000000000252. PMID: 22275487 Free PMC article. Retracted.
References
-
- Albers KM, Setzer RW, Taichman LB (1986) Heterogeneity in the replicating population of cultured human epidermal keratinocytes. Differentiation 31, 134. - PubMed
-
- Appleton DR, Wright NA, Dyson P (1977) The age distribution of cells in stratified squamous epithelium. J. Theor. Biol. 65, 769. - PubMed
-
- Bursch W, Kleine K, Tenniswood M (1990) The biochemistry of cell death by apoptosis. Biochem. Cell Biol. 68, 1071. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
