

Palmitic and stearic fatty acids induce caspase-dependent and -independent cell death in nerve growth factor differentiated PC12 cells
- PMID: 12562510
- PMCID: PMC4157900
- DOI: 10.1046/j.1471-4159.2003.01571.x
Palmitic and stearic fatty acids induce caspase-dependent and -independent cell death in nerve growth factor differentiated PC12 cells
Abstract
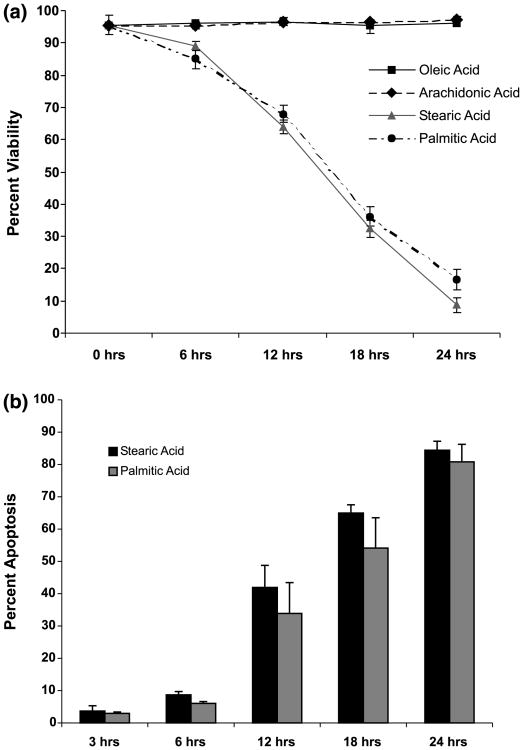
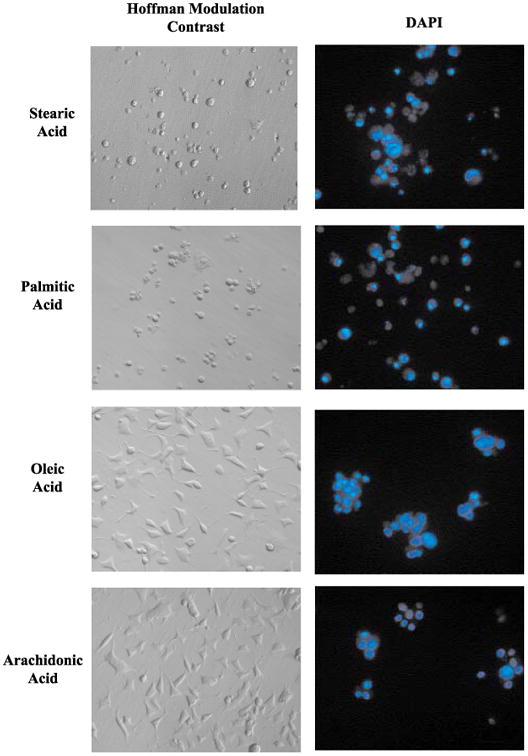
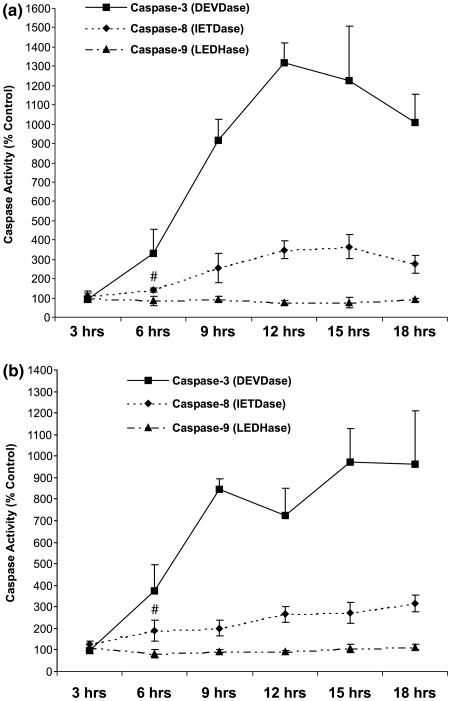
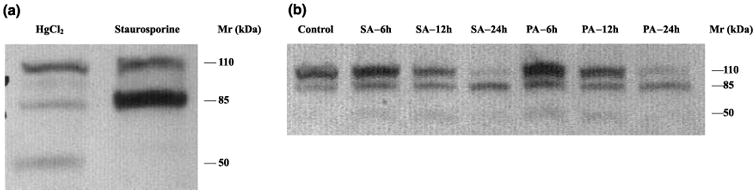
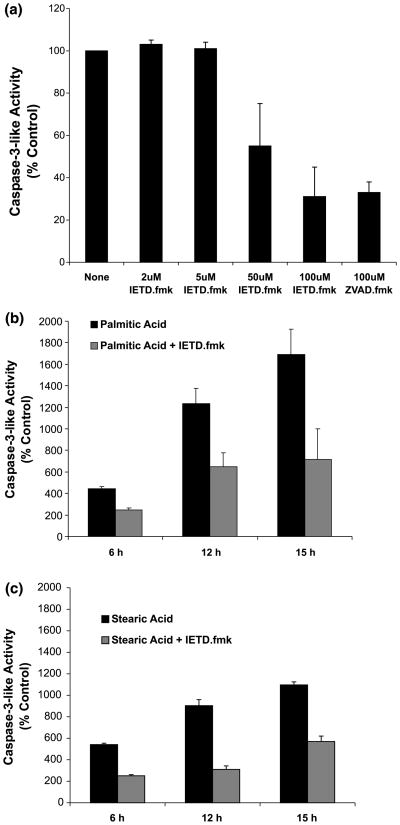
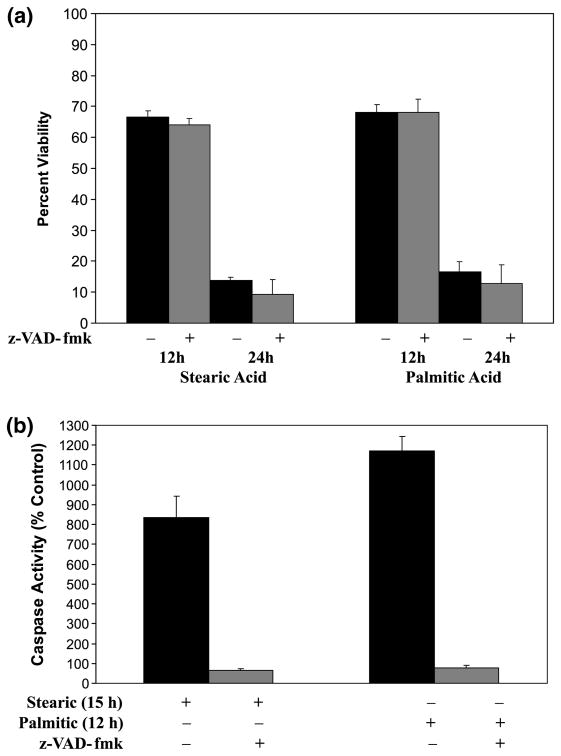
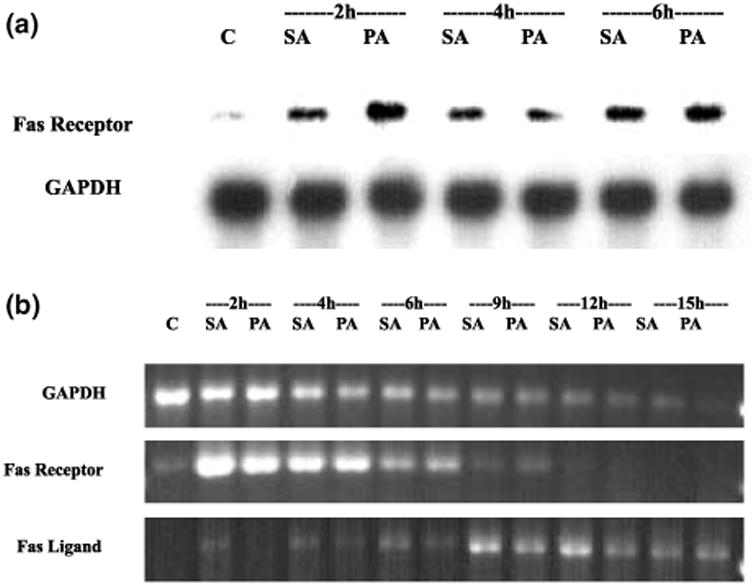
Apoptotic cell death has been proposed to play a role in the neuronal loss observed following traumatic injury in the CNS and PNS. The present study uses an in vitro tissue culture model to investigate whether free fatty acids (FFAs), at concentrations comparable to those found following traumatic brain injury, trigger cell death. Nerve growth factor (NGF)-differentiated PC12 cells exposed to oleic and arachidonic acids (2 : 1 ratio FFA/BSA) showed normal cell survival. However, when cells were exposed to stearic and palmitic acids, there was a dramatic loss of cell viability after 24 h of treatment. The cell death induced by stearic acid and palmitic acid was apoptotic as assessed by morphological analysis, and activation of caspase-8 and caspase-3-like activities. Western blotting showed that differentiated PC12 cells exposed to stearic and palmitic acids exhibited the signature apoptotic cleavage fragment of poly (ADP-ribose) polymerase (PARP). Interestingly, blockade of caspase activities with the pan-caspase inhibitor z-VAD-fmk failed to prevent the cell death observed induced by palmitic or stearic acid. RT-PCR and RNA blot experiments showed an up-regulation of the Fas receptor and ligand mRNA. These findings are consistent with our hypothesis that FFAs may play a role in the cell death associated with trauma in the CNS and PNS.
Figures
References
-
- Abe K, Kogure K, Yamamoto H, Imazawa M, Miyamoto K. Mechanism of arachidonic acid liberation during ischemia in gerbil cerebral cortex. J Neurochem. 1987;48:503–509. - PubMed
-
- Abe K, Yoshidomi M, Kogure K. Arachidonic acid metabolism in ischemic neuronal damage. Ann NY Acad Sci. 1991;559:259–268. - PubMed
-
- Bazan NG. Effects of ischemia and electroconvulsive shock on free fatty acid pool in the brain. Biochim Biophys Acta. 1970;218:1–10. - PubMed
-
- Bazan NG, De Bazan HEP, Kennedy WG, Joel CD. Regional distribution and rate of production of free fatty acids in rat brain. J Neurochem. 1971;18:1487–1393. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
