

The basic helix-loop-helix differentiation factor Nex1/MATH-2 functions as a key activator of the GAP-43 gene
- PMID: 12562512
- PMCID: PMC1413589
- DOI: 10.1046/j.1471-4159.2003.01572.x
The basic helix-loop-helix differentiation factor Nex1/MATH-2 functions as a key activator of the GAP-43 gene
Abstract
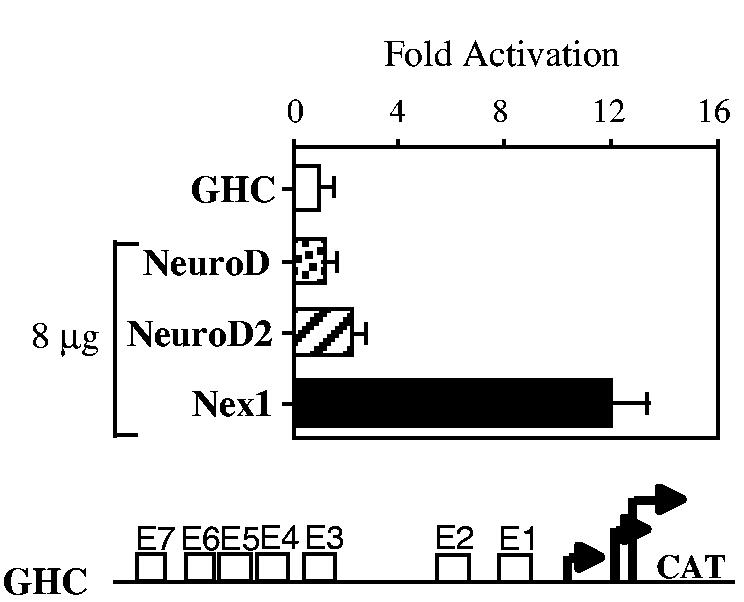
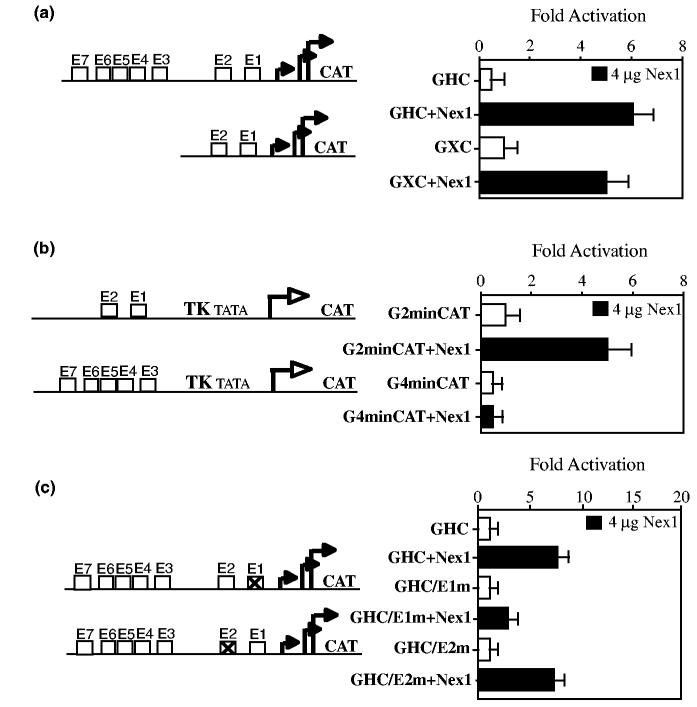
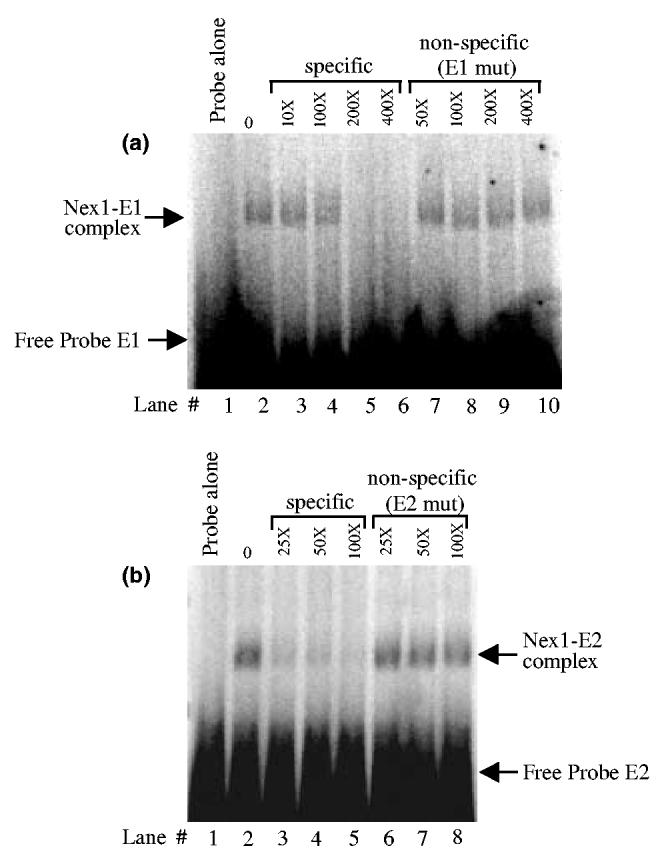
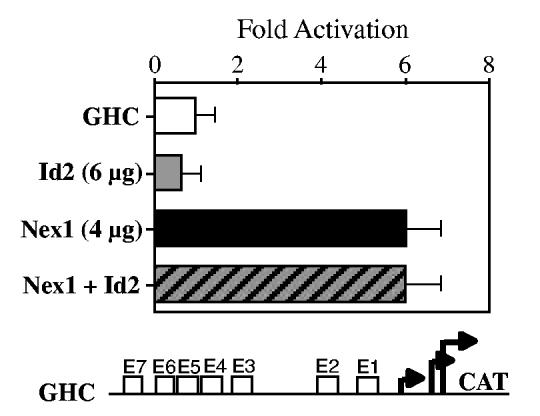
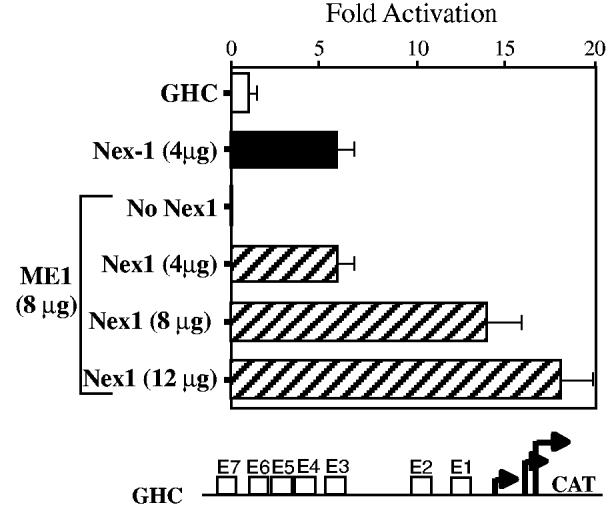
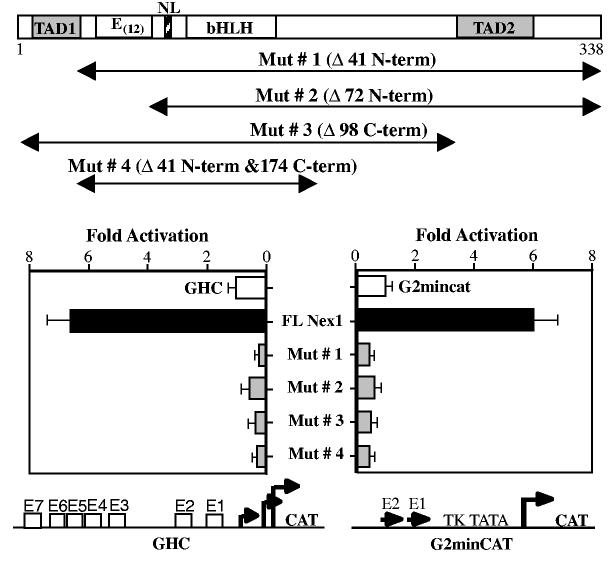
Nex1/MATH-2 is a neurogenic basic Helix-Loop-Helix (bHLH) transcription factor that belongs to the NeuroD subfamily. Its expression parallels that of the GAP-43 gene and peaks during brain development, when neurite outgrowth and synaptogenesis are highly active. We previously observed a direct correlation between the levels of expression of Nex1 and GAP-43 proteins, which resulted in extensive neurite outgrowth and neuronal differentiation of PC12 cells in the absence of nerve growth factor. Since the GAP-43 gene is a target for bHLH regulation, we investigated whether Nex1 could regulate the activity of the GAP-43 promoter. We found that among the members of the NeuroD subfamily, Nex1 promoted maximal activity of the GAP-43 promoter. The Nex1-mediated activity is restricted to the conserved E1-E2 cluster located near the major transcription start sites. By electrophoretic mobility shift assay and site-directed mutagenesis, we showed that Nex1 binds as homodimers and that the E1 E-box is a high affinity binding site. We further found that Nex1 released the ME1 E-protein-mediated repression in a concentration dependent manner. Thus, the E1-E2 cluster has a dual function: it can mediate activation or repression depending on the interacting bHLH proteins. Finally, a series of N-terminal and C-terminal deletions revealed that Nex1 transcriptional activity is linked to two distinct transactivation domains, TAD1 and TAD2, with TAD1 being unique to Nex1. Together, our results suggest that Nex1 may engage in selective interactions with components of the core transcriptional machinery whose assembly is dictated by the architecture of the GAP-43 promoter and cellular environment.
Figures
Similar articles
-
5'UTR of the neurogenic bHLH Nex1/MATH-2/NeuroD6 gene is regulated by two distinct promoters through CRE and C/EBP binding sites.J Neurosci Res. 2007 Jan;85(1):1-18. doi: 10.1002/jnr.21093. J Neurosci Res. 2007. PMID: 17075921 Free PMC article.
-
Constitutive overexpression of the basic helix-loop-helix Nex1/MATH-2 transcription factor promotes neuronal differentiation of PC12 cells and neurite regeneration.J Neurosci Res. 2002 Jan 15;67(2):235-45. doi: 10.1002/jnr.10119. J Neurosci Res. 2002. PMID: 11782967 Free PMC article.
-
The GAP-43 gene is a direct downstream target of the basic helix-loop-helix transcription factors.J Biol Chem. 1996 Sep 6;271(36):22035-43. doi: 10.1074/jbc.271.36.22035. J Biol Chem. 1996. PMID: 8703010
-
Vertebrate hairy and Enhancer of split related proteins: transcriptional repressors regulating cellular differentiation and embryonic patterning.Oncogene. 2001 Dec 20;20(58):8342-57. doi: 10.1038/sj.onc.1205094. Oncogene. 2001. PMID: 11840327 Review.
-
Basic helix-loop-helix transcription factors, BHLHB2 and BHLHB3; their gene expressions are regulated by multiple extracellular stimuli.Front Biosci. 2005 Sep 1;10:3151-71. doi: 10.2741/1772. Front Biosci. 2005. PMID: 15970569 Review.
Cited by
-
5'UTR of the neurogenic bHLH Nex1/MATH-2/NeuroD6 gene is regulated by two distinct promoters through CRE and C/EBP binding sites.J Neurosci Res. 2007 Jan;85(1):1-18. doi: 10.1002/jnr.21093. J Neurosci Res. 2007. PMID: 17075921 Free PMC article.
-
NeuroD6 genomic signature bridging neuronal differentiation to survival via the molecular chaperone network.J Neurosci Res. 2010 Jan;88(1):33-54. doi: 10.1002/jnr.22182. J Neurosci Res. 2010. PMID: 19610105 Free PMC article.
-
RNA-Seq data mining: downregulation of NeuroD6 serves as a possible biomarker for alzheimer's disease brains.Dis Markers. 2014;2014:123165. doi: 10.1155/2014/123165. Epub 2014 Dec 8. Dis Markers. 2014. PMID: 25548427 Free PMC article.
-
Math1 target genes are enriched with evolutionarily conserved clustered E-box binding sites.J Mol Neurosci. 2006;28(2):211-29. doi: 10.1385/JMN:28:2:211. J Mol Neurosci. 2006. PMID: 16679559
-
Homodimeric and Heterodimeric Interactions among Vertebrate Basic Helix-Loop-Helix Transcription Factors.Int J Mol Sci. 2021 Nov 28;22(23):12855. doi: 10.3390/ijms222312855. Int J Mol Sci. 2021. PMID: 34884664 Free PMC article. Review.
References
-
- Aigner L, Arber S, Kapfhammer JP, Laux T, Schneider C, Botteri F, Brenner HR, Caroni P. Overexpression of the neural growth-associated protein GAP-43 induces nerve sprouting in the adult nervous system of transgenic mice. Cell. 1995;83:269–278. - PubMed
-
- Anderson KD, Morin MA, Beckel-Mitchener A, Mobarak CD, Neve RL, Furneaux HM, Burry R, Perrone-Bizzozero NI. Overexpression of HuD, but not of its truncated form HuD I + II, promotes GAP-43 gene expression and neurite outgrowth in PC12 cells in the absence of nerve growth factor. J. Neurochem. 2000;75:1103–1114. - PubMed
-
- Bartholoma A, Nave KA. NEX-1: a novel brain-specific helix-loop-helix protein with autoregulation and sustained expression in mature cortical neurons. Mech. Dev. 1994;48:217–228. - PubMed
-
- Basi GS, Jacobson RD, Virag I, Schilling J, Skene JH. Primary structure and transcriptional regulation of GAP-43, a protein associated with nerve growth. Cell. 1987;49:785–791. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
