

Review
doi: 10.1128/JB.185.4.1128-1146.2003.
Prokaryotic development: emerging insights
Affiliations
- PMID: 12562781
- PMCID: PMC142891
- DOI: 10.1128/JB.185.4.1128-1146.2003
Item in Clipboard
Review
Prokaryotic development: emerging insights
J Bacteriol.
2003 Feb.
No abstract available
Figures
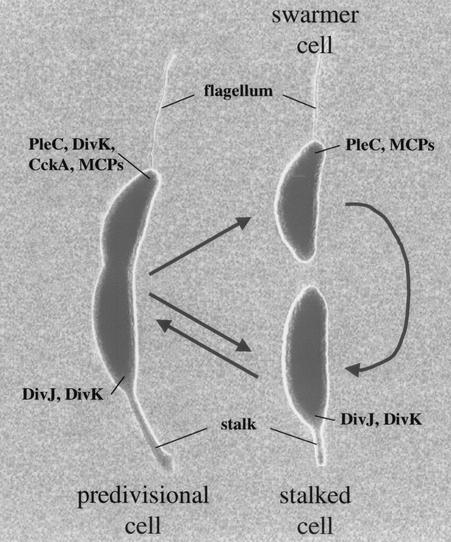
The C. crescentus cell cycle. The asymmetric predivisional cell divides to produce two distinct progeny: the flagellated swarmer cell and the sessile stalked cell. The stalked cell is competent to initiate DNA replication and cell growth to become a predivisional cell. The swarmer cell, however, first differentiates into a stalked cell. In addition to the external polar structures (stalk and flagellum), many membrane and membrane-associated proteins are found at specific cell poles during the cell cycle; some of these proteins are labeled.
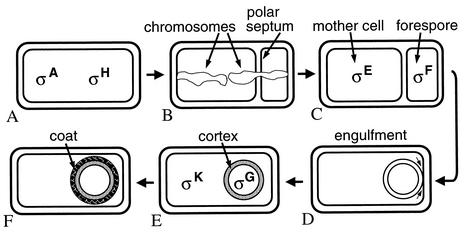
Morphological changes during B. subtilis sporulation and the approximate time and location at which different σ factors become active. (A) σA and σH RNA polymerases transcribe genes whose products cause polar septation and axial filament formation. (B) The axial filament consists of two chromosomes extending the length of the cell with their replication origin-proximal regions attached at opposite ends of the cell. The polar septum forms around the axial filament, capturing one-third of one chromosome in the forespore. The remaining two-thirds of that chromosome is translocated into the forespore. (C) Upon completion of polar septation, σF becomes active in the forespore, and this leads to activation of σE in the mother cell. (D) Products of genes under σE control drive migration of the septal membranes around the forespore in the phagocytic-like process of engulfment. (E) Completion of engulfment pinches off the forespore as a free protoplast within the mother cell. Two membranes surround the forespore and separate its contents from the mother cell cytoplasm. σG becomes active in the forespore, leading to activation of σK in the mother cell. Primarily, genes under σE and σK control cause synthesis of a loosely cross-linked peptidoglycan termed cortex, between the two membranes surrounding the forespore, and synthesis of proteins that assemble on the surface of the forespore to produce the spore coat (F). Not shown are subsequent steps, which include spore maturation and release of the spore via lysis of the mother cell. Adapted from reference with permission.
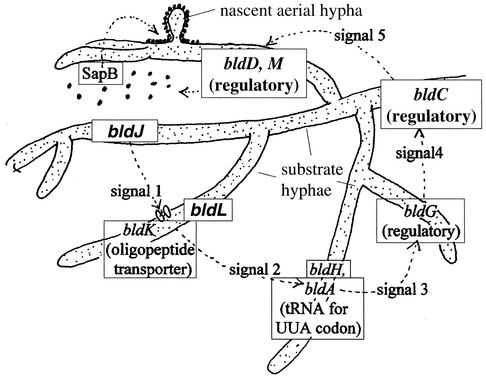
Extracellular signal exchange leading to aerial growth in S. coelicolor. The figure, based on data of Willey et al. (121) and Molle and Buttner (78), is modified from the work of Chater (16) with permission. The lower part depicts how substrate hyphae in different physiological states under bld gene control may produce signals that act in a cascade leading to production of SapB, which is thought to allow formation of aerial hyphae (one is illustrated near the top of the figure).
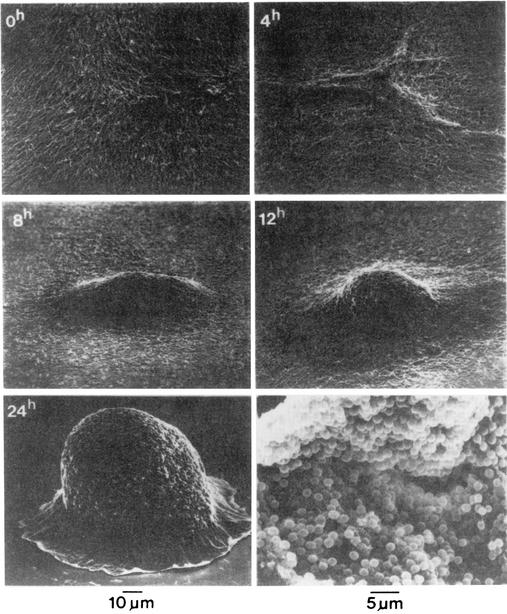
Fruiting body development in M. xanthus. Scanning electron micrographs of cells starved in submerged culture were made by Kuner and Kaiser (69) and are reprinted here with permission. Times poststarvation are indicated in the upper left corner of the first five frames, to which the 10-μm scale applies. Individual cells (about 5 μm long) begin to aggregate by 4 h and complete mound formation by 24 h, at which time some cells have begun to differentiate into spores. The lower right frame shows a mature fruiting body that has cracked open, revealing its spores (5-μm scale).
Similar articles
-
Modulation of bacterial proliferation as a survival strategy.Adv Appl Microbiol. 2015;92:127-71. doi: 10.1016/bs.aambs.2015.02.004. Epub 2015 Mar 25. Adv Appl Microbiol. 2015. PMID: 26003935 Review.
-
Wind River Conference on Prokaryotic Biology--2002.J Bacteriol. 2003 Jan;185(1):7-12. doi: 10.1128/JB.185.1.7-12.2003. J Bacteriol. 2003. PMID: 12486034 Free PMC article. No abstract available.
-
Cell regulation.Curr Opin Microbiol. 2012 Apr;15(2):115-7. doi: 10.1016/j.mib.2012.02.004. Epub 2012 Mar 10. Curr Opin Microbiol. 2012. PMID: 22410109 No abstract available.
-
You Are What You Eat: Metabolic Control of Bacterial Division.Trends Microbiol. 2016 Mar;24(3):181-189. doi: 10.1016/j.tim.2015.11.007. Epub 2015 Dec 12. Trends Microbiol. 2016. PMID: 26690613 Review.
-
Good things come in small packages: subcellular organization and development in bacteria.Curr Opin Microbiol. 2011 Dec;14(6):687-90. doi: 10.1016/j.mib.2011.10.011. Epub 2011 Nov 14. Curr Opin Microbiol. 2011. PMID: 22088394 No abstract available.
Cited by
-
Lmo0171, a novel internalin-like protein, determines cell morphology of Listeria monocytogenes and its ability to invade human cell lines.Curr Microbiol. 2015 Feb;70(2):267-74. doi: 10.1007/s00284-014-0715-4. Epub 2014 Oct 17. Curr Microbiol. 2015. PMID: 25323012 Free PMC article.
-
patS minigenes inhibit heterocyst development of Anabaena sp. strain PCC 7120.J Bacteriol. 2004 Oct;186(19):6422-9. doi: 10.1128/JB.186.19.6422-6429.2004. J Bacteriol. 2004. PMID: 15375122 Free PMC article.
-
Rhizosheath microbial community assembly of sympatric desert speargrasses is independent of the plant host.Microbiome. 2018 Dec 4;6(1):215. doi: 10.1186/s40168-018-0597-y. Microbiome. 2018. PMID: 30514367 Free PMC article.
-
Helical disposition of proteins and lipopolysaccharide in the outer membrane of Escherichia coli.J Bacteriol. 2005 Mar;187(6):1913-22. doi: 10.1128/JB.187.6.1913-1922.2005. J Bacteriol. 2005. PMID: 15743937 Free PMC article.
-
Transcriptional Profiling of Coxiella burnetii Reveals Extensive Cell Wall Remodeling in the Small Cell Variant Developmental Form.PLoS One. 2016 Feb 24;11(2):e0149957. doi: 10.1371/journal.pone.0149957. eCollection 2016. PLoS One. 2016. PMID: 26909555 Free PMC article.
References
-
- Adams, D. G. 2000. Heterocyst formation in cyanobacteria. Curr. Opin. Microbiol. 3:618-624. - PubMed
-
- Aldridge, P., and K. T. Hughes. 2002. Regulation of flagellar assembly. Curr. Opin. Microbiol. 5:160-165. - PubMed
-
- Alley, M. R., J. R. Maddock, and L. Shapiro. 1993. Requirement of the carboxyl terminus of a bacterial chemoreceptor for its targeted proteolysis. Science 259:1754-1757. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
