

Branching of Escherichia coli cells arises from multiple sites of inert peptidoglycan
- PMID: 12562782
- PMCID: PMC142844
- DOI: 10.1128/JB.185.4.1147-1152.2003
Branching of Escherichia coli cells arises from multiple sites of inert peptidoglycan
Abstract
Some strains of Escherichia coli defective for dacA, the gene coding for penicillin-binding protein 5, exhibit a strong branching phenotype when cell division is blocked. Since such branch formation implies a differentiation of polar caps at ectopic locations in the cell envelope, we analyzed murein segregation and observed a strong correlation between areas of inert murein and these morphological anomalies. In particular, the tips of branches exhibited the same properties as those described for polar caps of wild-type cells, i.e., the synthesis and turnover of murein were inhibited. Also, the mobility of cell envelope proteins was apparently constrained in areas with morphological defects. Polar regions of branching cells and sacculi had aberrant morphologies with a very high frequency. Of special interest was that areas of inert murein at polar caps were often split by areas of active synthesis, a situation unlike that observed in wild-type cells. These observations suggest that in dacA mutants, branches and other morphological anomalies may arise from split polar caps or by de novo generation of new poles built around inert peptidoglycan patches in the side walls of the cell.
Figures
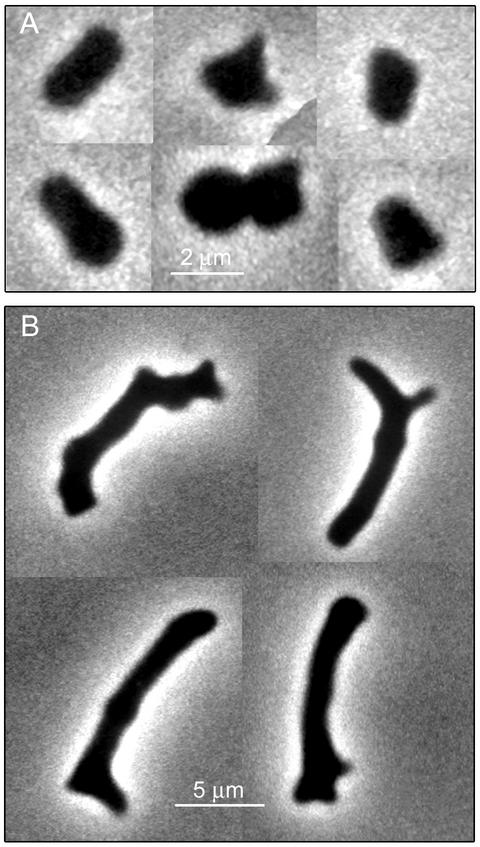
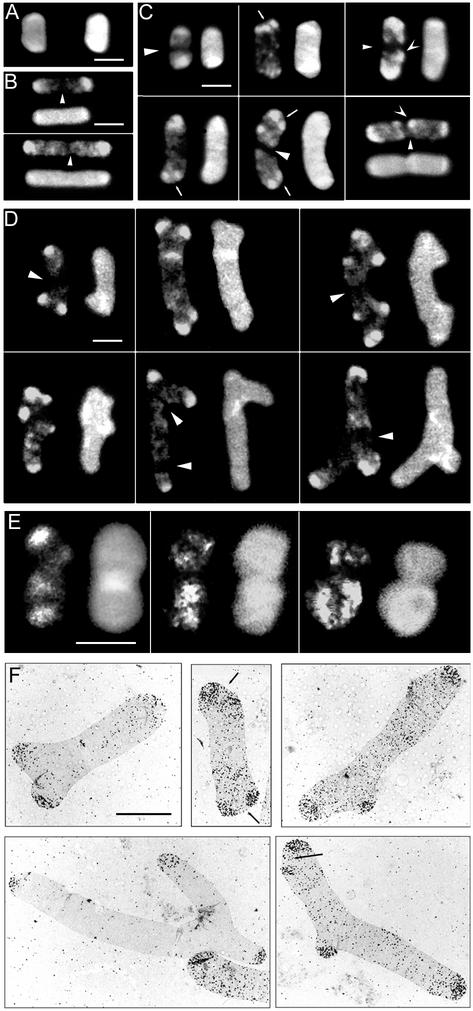
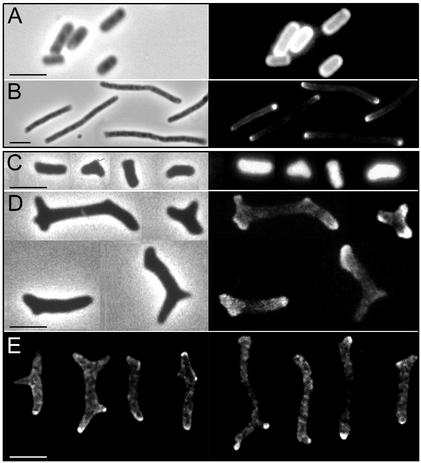
Comment in
-
New insights into the developmental history of the bacterial cell division site.J Bacteriol. 2003 Feb;185(4):1125-7. doi: 10.1128/JB.185.4.1125-1127.2003. J Bacteriol. 2003. PMID: 12562780 Free PMC article. No abstract available.
References
-
- Ayala, J. A., T. Garrido, M. A. de Pedro, and M. Vicente. 1994. Molecular biology of bacterial septation, p. 73-101. In J. M. Ghuysen and R. Hakenbeck (ed.), Bacterial cell wall. Elsevier, Amsterdam, The Netherlands.
-
- Cooper, S. 1991. Bacterial growth and division. Academic Press, Inc., San Diego, Calif.
-
- Danscher, G. 1981. Histochemical demonstration of heavy metals. A revised version of the sulphide silver method suitable for both light and electron microscopy. Histochemistry 71:1-16. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
