

doi: 10.1128/JB.185.4.1470-1474.2003.
Overrepresentation of a gene family encoding extracytoplasmic solute receptors in Bordetella
Affiliations
- PMID: 12562821
- PMCID: PMC142875
- DOI: 10.1128/JB.185.4.1470-1474.2003
Item in Clipboard
Overrepresentation of a gene family encoding extracytoplasmic solute receptors in Bordetella
J Bacteriol.
2003 Feb.
Abstract
A family of genes that are likely to encode extracytoplasmic solute receptors is strongly overrepresented in several beta-proteobacteria, including Bordetella pertussis. This gene family, of which members have been called bug genes, contains some examples that are contained within polycistronic operons coding for tripartite uptake transporters of the TTT family, while the vast majority are "orphan" genes. Proteomic and functional analyses demonstrated that several of these genes are expressed in B. pertussis, and one is involved in citrate uptake. The bug genes probably form an ancient family that has been subjected to a large expansion in a restricted phylogenic group.
Figures
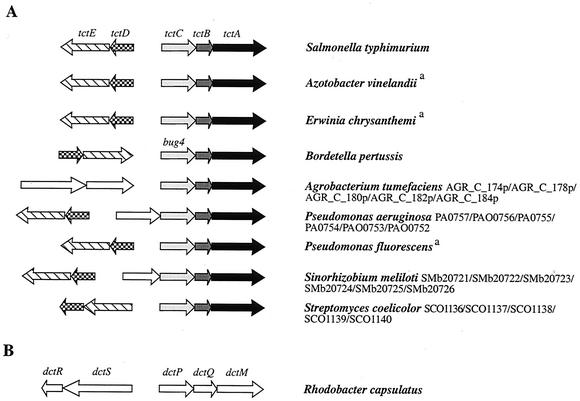
Organization of selected operons encoding predicted TTTs and comparison with the dctPQM operon that encodes the archetype of TRAP transporters. (A) tct operon of S. enterica serovar Typhimurium is shown as the paradigm for TTT systems. Homologs of tctC, tctB, and tctA are shown in light gray, dark gray, and black, respectively. Homologs of tctE and tctD, coding for the sensor kinase and response regulator of a two-component system involved in the regulation of tctCBA (29), are depicted as hatched and checkered arrows, respectively. Among the three predicted TTT-coding operons of Agrobacterium tumefaciens, only the one depicted here is preceded by an operon encoding a two-component system. It is not homologous to TctDE but to DctRS of Rhodobacter capsulatus, involved in the regulation of the dicarboxylate transporter operon dctPQM (see panel B). Additional genes (in white) are found in a few operons; these include genes presumably coding for a porin in Pseudomonas aeruginosa and for an ESR associated with ABC uptake transporters in Sinorhizobium meliloti. a, no accession numbers are yet available for Azotobacter vinelandii, Erwinia chrysanthemi, and Pseudomonas fluorescens. (B) The dctPQM operon that encodes the dicarboxylate TRAP transporter of Rhodobacter capsulatus (10) and the adjacent dctRS operon that encodes a two-component system are shown for comparison purposes. Although not homologous, TTT and TRAP transporters have similar molecular organizations.
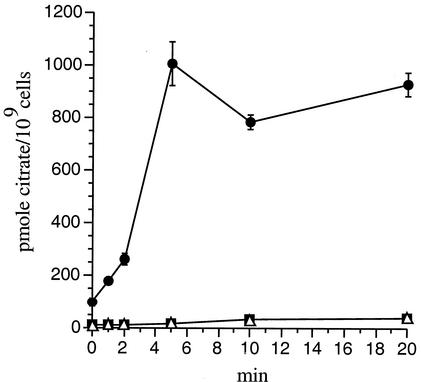
Citrate uptake by B. pertussis strains. BPSM carrying an intact copy of the operon (black circles) and two isogenic mutants, one with an in-frame deletion of the bug4 gene (black squares) and the other carrying a knockout mutation of the B. pertussis tctA ortholog (open triangles), were used to monitor the uptake of radiolabeled citrate over time. The results are expressed as picomoles of radioactive citrate incorporated by 109 cells.
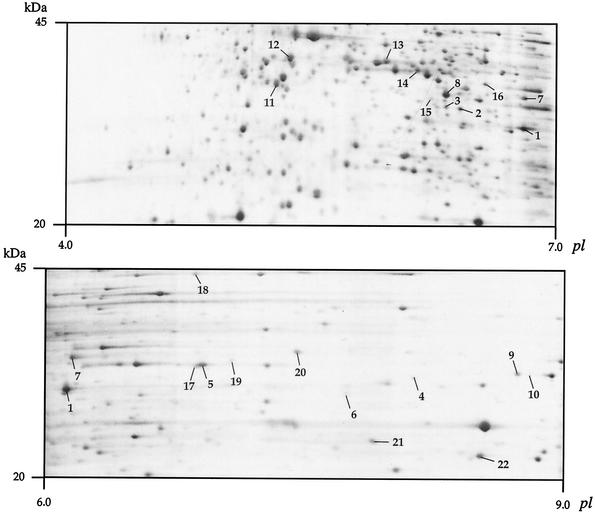
Analyses of B. pertussis BPDR proteins by 2-D electrophoresis. The portions of pH 4 to 7 (top) and pH 6 to 9 (bottom) silver-stained gels corresponding to the 20- to 45-kDa range are shown. Only the putative periplasmic proteins involved in solute binding as identified by mass fingerprinting have been numbered (1 to 6, Bug9, -2, -73, -20, -71, and -4, respectively; 7 to 10, predicted TRAP transporter-associated ESRs; 11 to 22, predicted ABC transporter-associated ESRs). See http://www.ibl.fr/articles/jbact_antoine.htm (Table A3) for the sequences of these 22 proteins.
References
-
- Antoine, R., S. Alonzo, D. Raze, L. Coutte, S. Lesjean, E. Willery, C. Locht, and F. Jacob-Dubuisson. 2000. New virulence-activated and virulence-repressed genes identified by systematic gene inactivation and generation of transcriptional fusions in Bordetella pertussis. J. Bacteriol. 182:5902-5905. - PMC - PubMed
-
- Antoine, R., D. Raze, and C. Locht. 2000. Genomics of Bordetella pertussis toxins. Int. J. Med. Microbiol. 290:301-305. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
