

Properties of the human muscle nicotinic receptor, and of the slow-channel myasthenic syndrome mutant epsilonL221F, inferred from maximum likelihood fits
- PMID: 12562900
- PMCID: PMC2342726
- DOI: 10.1113/jphysiol.2002.034173
Properties of the human muscle nicotinic receptor, and of the slow-channel myasthenic syndrome mutant epsilonL221F, inferred from maximum likelihood fits
Abstract
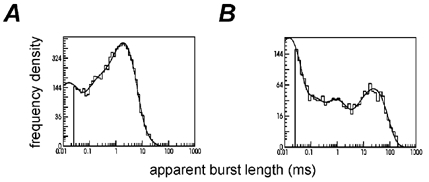
The mechanisms that underlie activation of nicotinic receptors are investigated using human recombinant receptors, both wild type and receptors that contain the slow channel myasthenic syndrome mutation, epsilonL221F. The method uses the program HJCFIT, which fits the rate constants in a specified mechanism directly to a sequence of observed open and shut times by maximising the likelihood of the sequence with exact correction for missed events. A mechanism with two different binding sites was used. The rate constants that apply to the diliganded receptor (opening, shutting and total dissociation rates) were estimated robustly, being insensitive to the exact assumptions made during fitting, as expected from simulation studies. They are sufficient to predict the main physiological properties of the receptors. The epsilonL221F mutation causes an approximately 4-fold reduction in dissociation rate from diliganded receptors, and a smaller increase in opening rate and mean open time. These are sufficient to explain the approximately 6-fold slowing of decay of miniature synaptic currents seen in patients. The distinction between the two binding sites was less robust, the estimates of rate constants being dependent to some extent on assumptions, e.g. whether an extra short-lived shut state was included or whether the EC50 was constrained. The results suggest that the two binding sites differ by roughly 10-fold in the affinity of the shut receptor for ACh in the wild type, and that in the epsilonL221F mutation the lower affinity is increased so the sites become more similar.
Figures













References
-
- Akabas M, Karlin A. Identification of acetylcholine receptor channel-lining residues in the M1 segment of the α-subunit. Biochemistry. 1995;34:12496–12500. - PubMed
-
- Beeson D, Brydson M, Betty M, Jeremiah S, Povey S, Vincent A, Newsom-Davis J. Primary structure of the human muscle acetylcholine receptor cDNA cloning of the τ and ε subunits. Eur J Biochem. 1993;215:229–238. - PubMed
-
- Beeson D, Newsom-Davis J. Mutations affecting muscle acetylcholine receptors and their role in congenital myasthenic syndromes. In: Lehmann-Horn F, editor. Channelopathies. BV: Elsevier Science; 2000. pp. 85–112.
MeSH terms
Substances
LinkOut - more resources
Full Text Sources