

The quality of maximum likelihood estimates of ion channel rate constants
- PMID: 12562901
- PMCID: PMC2342730
- DOI: 10.1113/jphysiol.2002.034165
The quality of maximum likelihood estimates of ion channel rate constants
Abstract
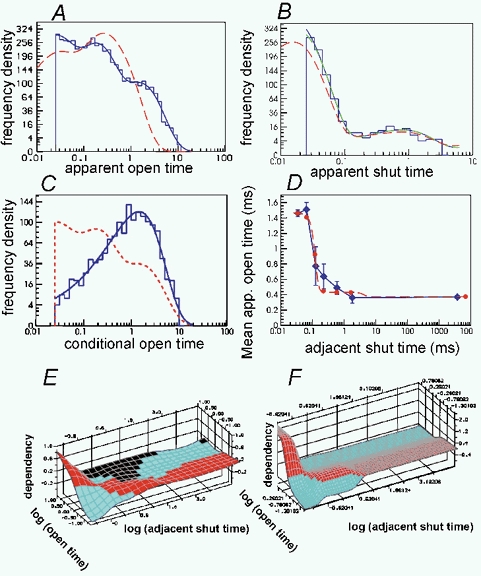
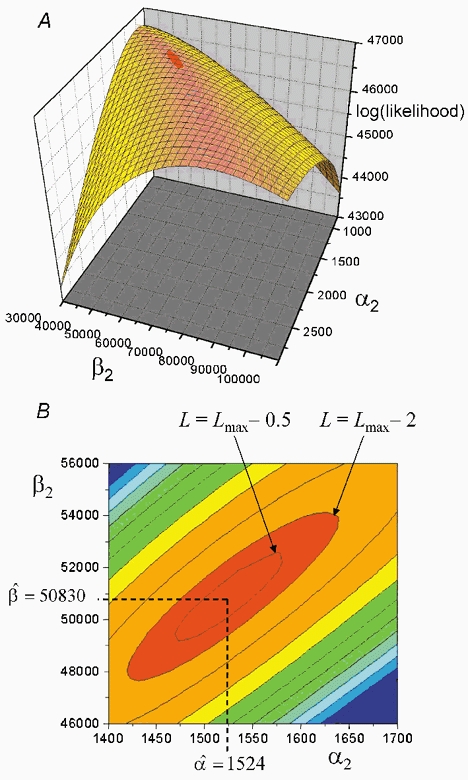
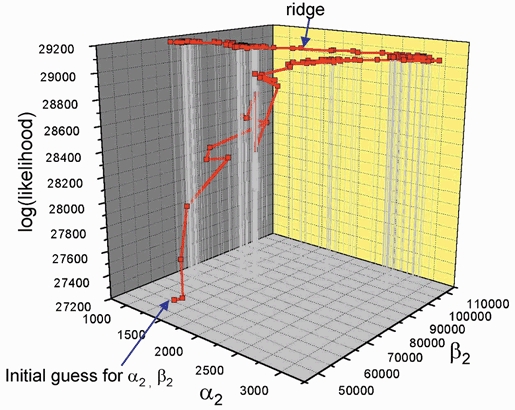
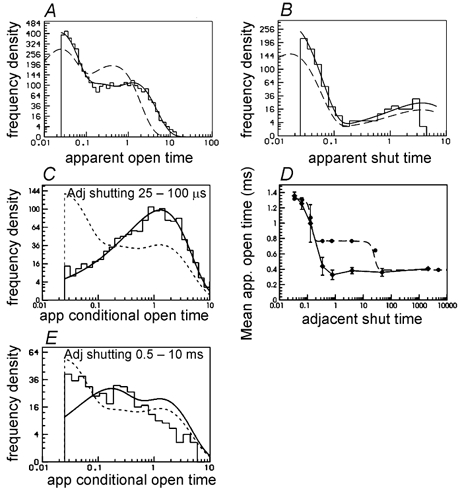
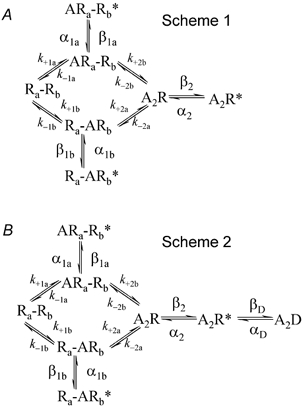
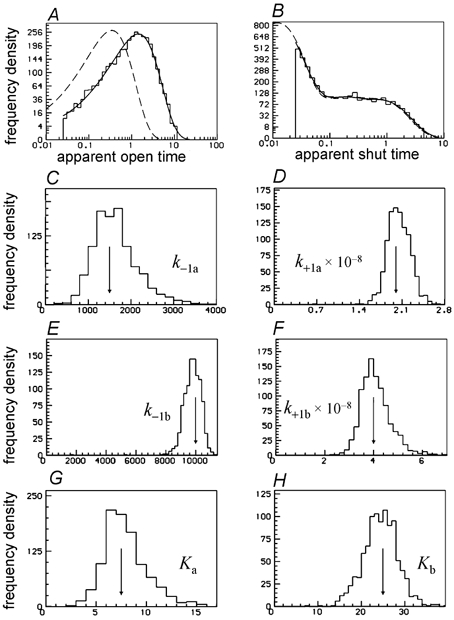
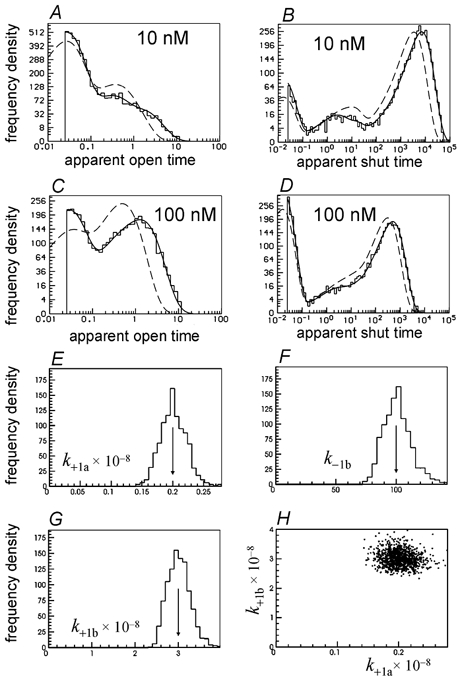
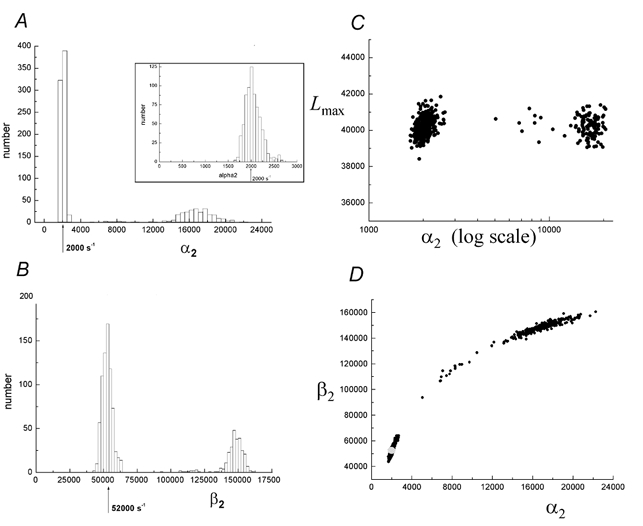
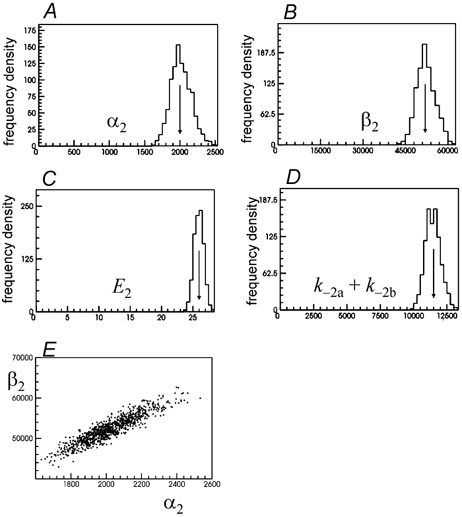
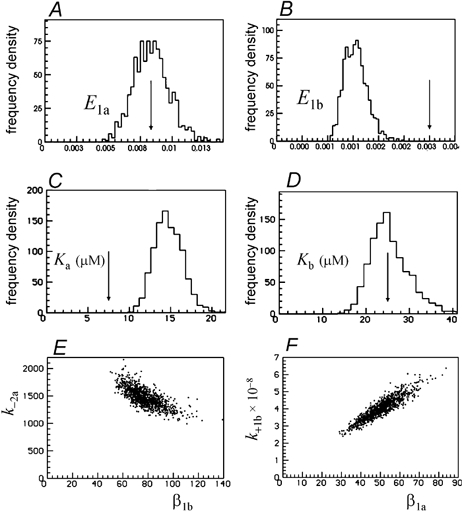
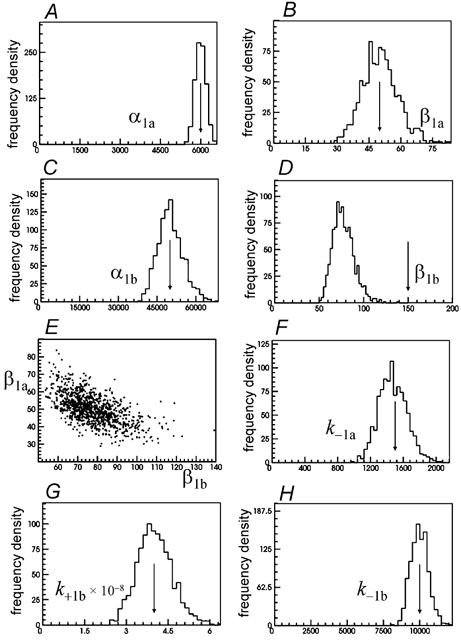
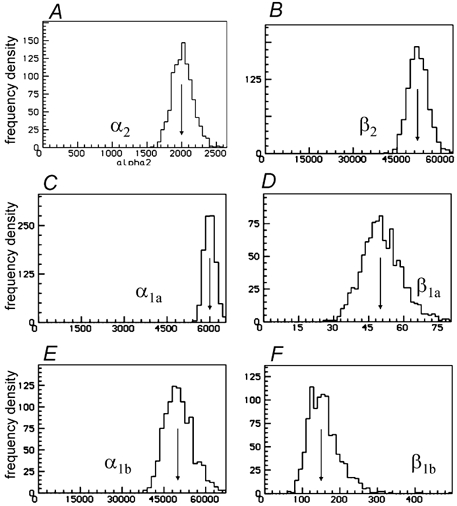
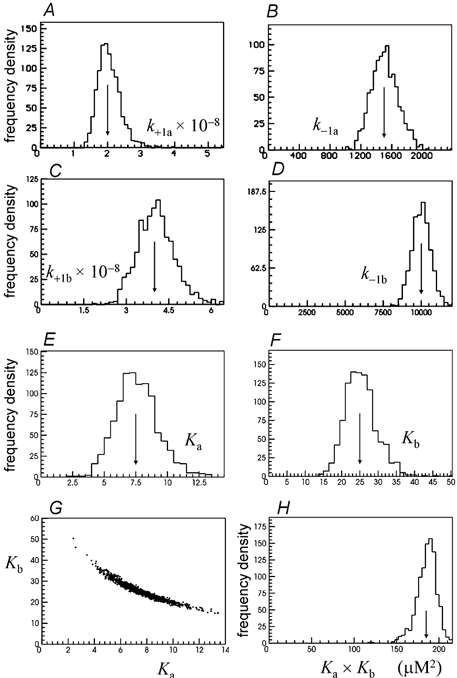
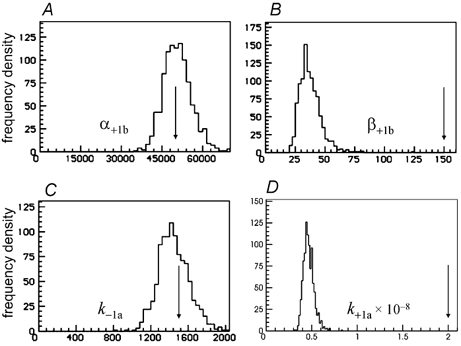
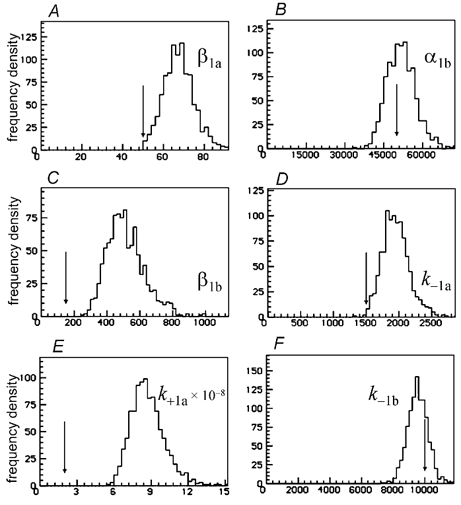
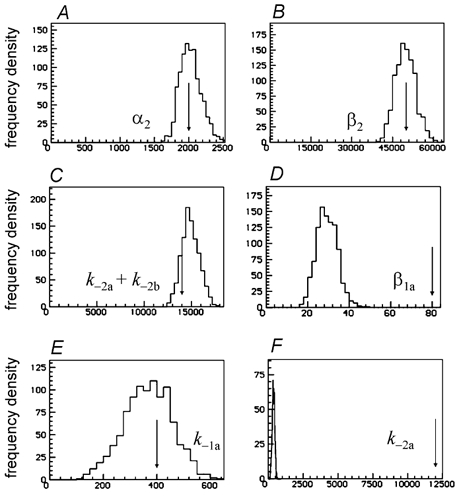
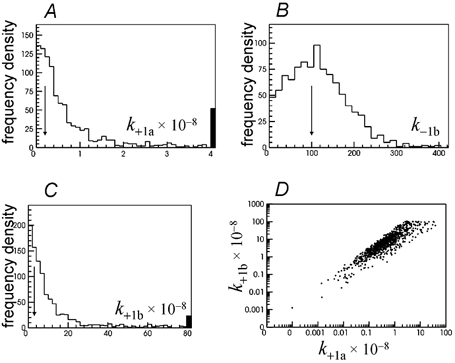
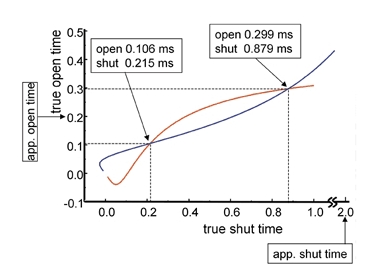
Properties of maximum likelihood estimators of rate constants for channel mechanisms are investigated, to see what can and cannot be inferred from experimental results. The implementation of the HJCFIT method is described; it maximises the likelihood of an entire sequence of apparent open and shut times, with the rate constants in a specified reaction mechanism as free parameters. The exact method for missed brief events is used. Several methods for testing the quality of the fit are described. The distributions of rate constants, and correlations between them, are investigated by doing sets of 1000 fits to simulated experiments. In a standard nicotinic receptor mechanism, all nine free rate constants can be estimated even from one single channel recording, as long as the two binding sites are independent, even when the number of channels in the patch is not known. The estimates of rate constants that apply to diliganded channels are robust; good estimates can be obtained even with erroneous assumptions (e.g. about the value of a fixed rate constant or the independence of sites). Rate constants that require distinction between the two sites are less robust, and require that an EC50 be specified, or that records at two concentrations be fitted simultaneously. Despite the complexity of the problem, it appears that there exist two solutions with very similar likelihoods, as in the simplest case. The hazards that result from this, and from the strong positive correlation between estimates of opening and shutting rates, are discussed.
Figures

References
-
- Ball FG, Davies SS, Sansom MSP. Aggregated Markov processes incorporating time interval omission. Adv Appl Prob. 1988a;20:546–572.
-
- Ball FG, Davies SS, Sansom MS. Single-channel data and missed events: analysis of a two-state Markov model. Proc R Soc Lond B Biol Sci. 1990;242:61–67. - PubMed
-
- Ball FG, Sansom MSP. Ion channel gating mechanisms: model identification and parameter estimation from single channel recordings. Proc R Soc Lond B Biol Sci. 1989;236:385–416. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources