

Enhanced G protein-dependent modulation of excitatory synaptic transmission in the cerebellum of the Ca2+ channel-mutant mouse, tottering
- PMID: 12562906
- PMCID: PMC2342657
- DOI: 10.1113/jphysiol.2002.033415
Enhanced G protein-dependent modulation of excitatory synaptic transmission in the cerebellum of the Ca2+ channel-mutant mouse, tottering
Abstract
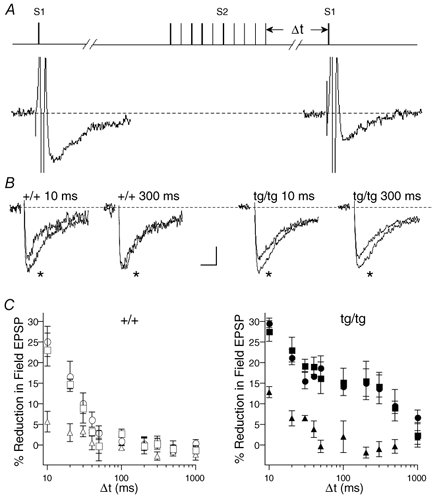
Tottering, a mouse model for absence epilepsy and cerebellar ataxia, carries a mutation in the gene encoding class A (P/Q-type) Ca2+ channels, the dominant exocytotic Ca2+ channel at most synapses in the mammalian central nervous system. Comparing tottering to wild-type mice, we have studied glutamatergic transmission between parallel fibres and Purkinje cells in cerebellar slices. Results from biochemical assays and electrical field recordings demonstrate that glutamate release from parallel fibre terminals of the tottering mouse is controlled largely by class B Ca2+ channels (N-type), in contrast to the P/Q-channels that dominate release from wild-type terminals. Since N-channels, in a variety of assays, are more effectively inhibited by G proteins than are P/Q-channels, we tested whether synaptic transmission between parallel fibres and Purkinje cells in tottering mice was more susceptible to inhibitory modulation by G protein-coupled receptors than in their wild-type counterparts. GABAB receptors and alpha2-adrenergic receptors (activated by bath application of transmitters) produced a three- to fivefold more potent inhibition of transmission in tottering than in wild-type synapses. This increased modulation is likely to be important for cerebellar transmission in vivo, since heterosynaptic depression, produced by activating GABAergic interneurones, greatly prolonged GABAB receptor-mediated presynaptic inhibition in tottering as compared to wild-type slices. We propose that this enhanced modulation shifts the balance of synaptic input to Purkinje cells in favour of inhibition, reducing Purkinje cell output from the cerebellum, and may contribute to the aberrant motor phenotype that is characteristic of this mutant animal.
Figures







References
-
- Abbott LC, Bump M, Brandl A, De Laune S. Investigation of the role of the cerebellum in the myoclonic-like movement disorder exhibited by tottering mice. Mov Disord. 2000;15:53–59. - PubMed
-
- Buchhalter JR. Animal models of inherited epilepsy. Epilepsia. 1993;3:S31–41. - PubMed
-
- Burgess DL, Noebels JL. Voltage-dependent calcium channel mutations in neurological disease. Ann NY Acad Sci. 1999;868:199–212. - PubMed
-
- Caddick SJ, Wang C, Fletcher CF, Jenkins NA, Copeland NG, Hosford DA. Excitatory but not inhibitory synaptic transmission is reduced in lethargic (Cacnb4(lh) and tottering (Cacna1atg) mouse thalami. J Neurophysiol. 1999;81:2066–2074. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous