

Cl- flux through a non-selective, stretch-sensitive conductance influences the outer hair cell motor of the guinea-pig
- PMID: 12562920
- PMCID: PMC2342734
- DOI: 10.1113/jphysiol.2002.036434
Cl- flux through a non-selective, stretch-sensitive conductance influences the outer hair cell motor of the guinea-pig
Abstract
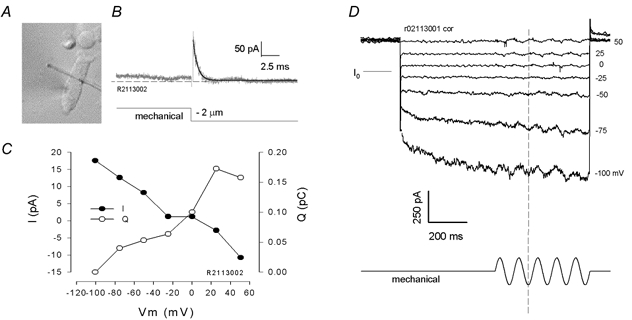
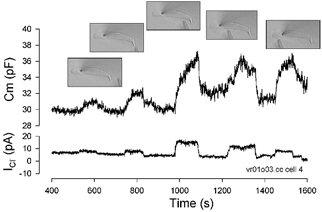
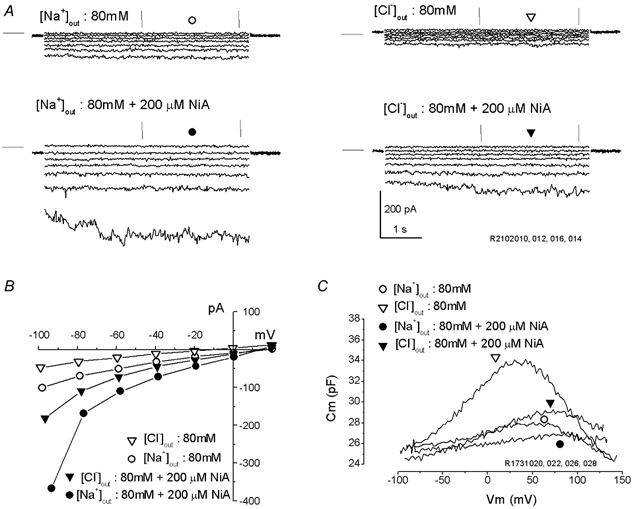
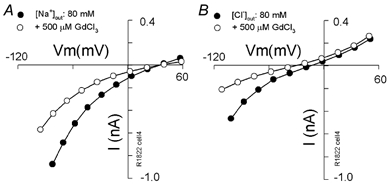
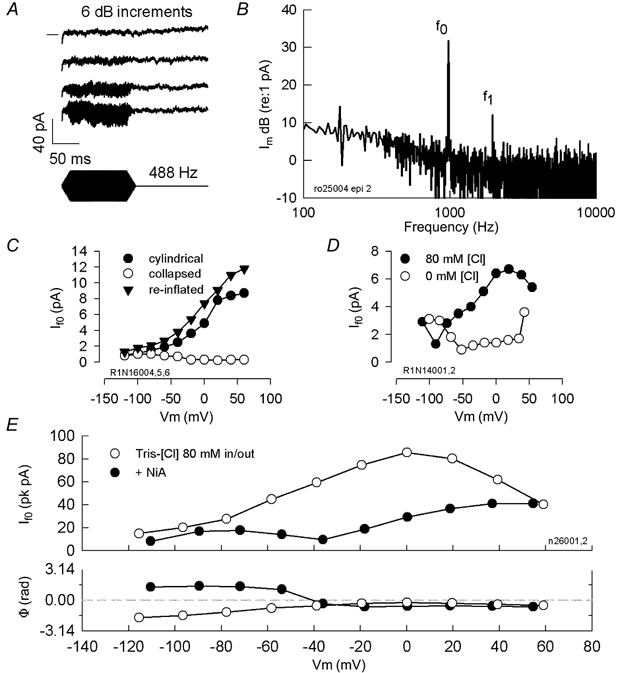
Outer hair cells underlie high frequency cochlear amplification in mammals. Fast somatic motility can be driven by voltage-dependent conformational changes in the motor protein, prestin, which resides exclusively within lateral plasma membrane of the cell. Yet, how a voltage-driven motor could contribute to high frequency amplification, despite the low-pass membrane filter of the cell, remains an enigma. The recent identification of prestin's Cl- sensitivity revealed an alternative mechanism in which intracellular Cl- fluctuations near prestin could influence the motor. We report the existence of a stretch-sensitive conductance within the lateral membrane that passes anions and cations and is gated at acoustic rates. The resultant intracellular Cl- oscillations near prestin may drive motor protein transitions, as evidenced by pronounced shifts in prestin's state-probability function along the voltage axis. The sensitivity of prestin's state probability to intracellular Cl- levels betokens a more complicated role for Cl- than a simple extrinsic voltage sensor. Instead, we suggest an allosteric modulation of prestin by Cl- and other anions. Finally, we hypothesize that prestin sensitivity to anion flux through the mechanically activated lateral membrane can provide a driving force that circumvents the membrane's low-pass filter, thus permitting amplification at high acoustic frequencies.
Figures








Similar articles
-
Chloride Anions Regulate Kinetics but Not Voltage-Sensor Qmax of the Solute Carrier SLC26a5.Biophys J. 2016 Jun 7;110(11):2551-2561. doi: 10.1016/j.bpj.2016.05.002. Biophys J. 2016. PMID: 27276272 Free PMC article.
-
Outer hair cell electromotility is low-pass filtered relative to the molecular conformational changes that produce nonlinear capacitance.J Gen Physiol. 2019 Dec 2;151(12):1369-1385. doi: 10.1085/jgp.201812280. Epub 2019 Nov 1. J Gen Physiol. 2019. PMID: 31676485 Free PMC article.
-
Conformational state-dependent anion binding in prestin: evidence for allosteric modulation.Biophys J. 2010 Feb 3;98(3):371-6. doi: 10.1016/j.bpj.2009.10.027. Biophys J. 2010. PMID: 20141749 Free PMC article.
-
Progress in understanding the structural mechanism underlying prestin's electromotile activity.Hear Res. 2022 Sep 15;423:108423. doi: 10.1016/j.heares.2021.108423. Epub 2021 Dec 24. Hear Res. 2022. PMID: 34987017 Review.
-
Prestin at year 14: progress and prospect.Hear Res. 2014 May;311:25-35. doi: 10.1016/j.heares.2013.12.002. Epub 2013 Dec 17. Hear Res. 2014. PMID: 24361298 Free PMC article. Review.
Cited by
-
On the temperature and tension dependence of the outer hair cell lateral membrane conductance GmetL and its relation to prestin.Pflugers Arch. 2006 Jun;452(3):283-9. doi: 10.1007/s00424-005-0037-2. Epub 2006 Mar 7. Pflugers Arch. 2006. PMID: 16521020
-
Prestin and the cochlear amplifier.J Physiol. 2006 Oct 1;576(Pt 1):37-42. doi: 10.1113/jphysiol.2006.114652. Epub 2006 Jul 27. J Physiol. 2006. PMID: 16873410 Free PMC article. Review.
-
The chloride-channel blocker 9-anthracenecarboxylic acid reduces the nonlinear capacitance of prestin-associated charge movement.Eur J Neurosci. 2016 Apr;43(8):1062-74. doi: 10.1111/ejn.13209. Epub 2016 Mar 16. Eur J Neurosci. 2016. PMID: 26869218 Free PMC article.
-
Making an effort to listen: mechanical amplification in the ear.Neuron. 2008 Aug 28;59(4):530-45. doi: 10.1016/j.neuron.2008.07.012. Neuron. 2008. PMID: 18760690 Free PMC article. Review.
-
Membrane prestin expression correlates with the magnitude of prestin-associated charge movement.Hear Res. 2016 Sep;339:50-9. doi: 10.1016/j.heares.2016.05.016. Epub 2016 Jun 1. Hear Res. 2016. PMID: 27262187 Free PMC article.
References
-
- Allen MC, Newland C, Valverde MA, Hardy SP. Inhibition of ligand-gated cation-selective channels by tamoxifen. Eur J Pharmacol. 1998;354:261–269. - PubMed
-
- Ashmore JF. The cellular physiology of isolated outer hair cells: implications for cochlear frequency selectivity. In: Moore BC, Patterson RD, editors. Auditory Frequency Selectivity. New York: Plenum Press; 1986. pp. 103–108.
-
- Ashmore JF. Transducer motor coupling in cochlear outer hair cells. In: Kemp D, Wilson JP, editors. Mechanics of Hearing. New York: Plenum Press; 1989. pp. 107–113.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous