

Mast cell tryptase and proteinase-activated receptor 2 induce hyperexcitability of guinea-pig submucosal neurons
- PMID: 12562962
- PMCID: PMC2342663
- DOI: 10.1113/jphysiol.2002.032011
Mast cell tryptase and proteinase-activated receptor 2 induce hyperexcitability of guinea-pig submucosal neurons
Abstract
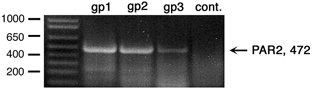
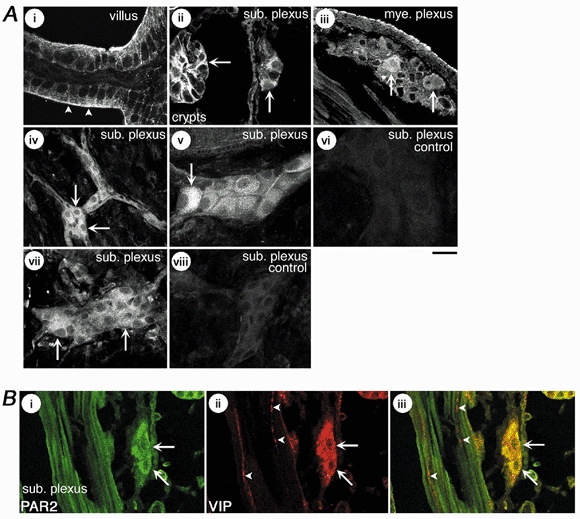
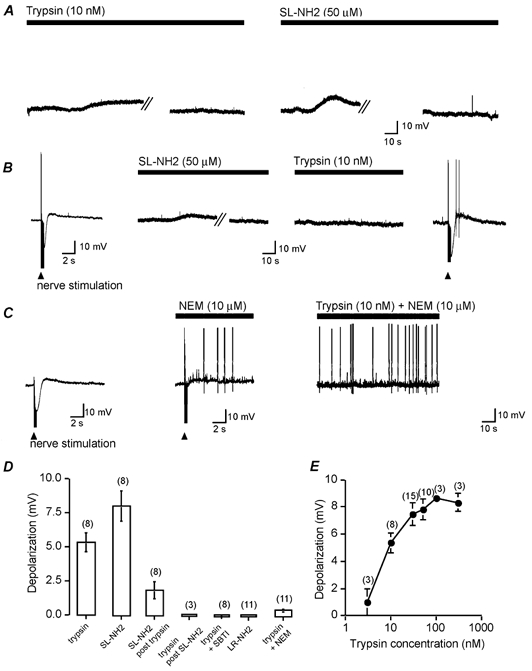
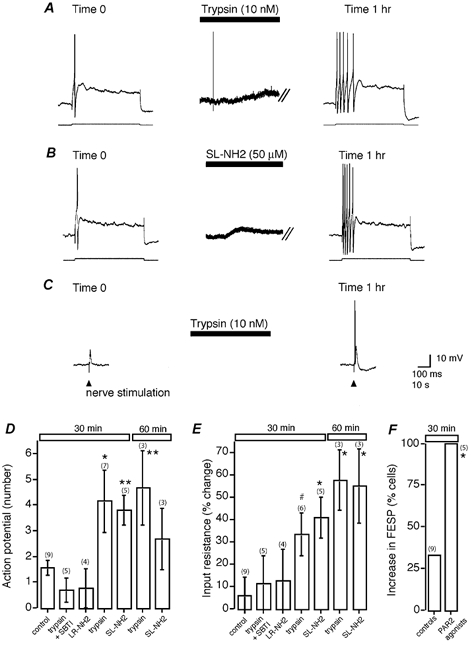
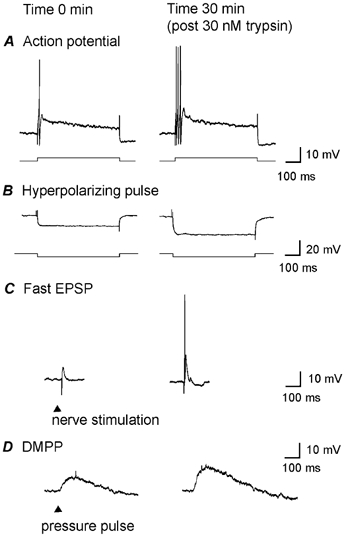
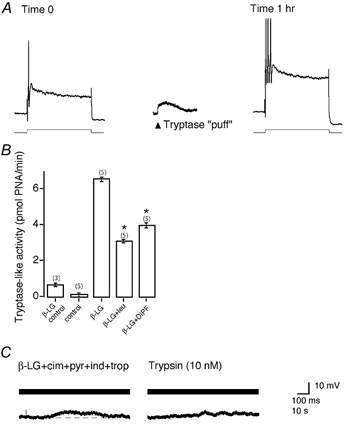
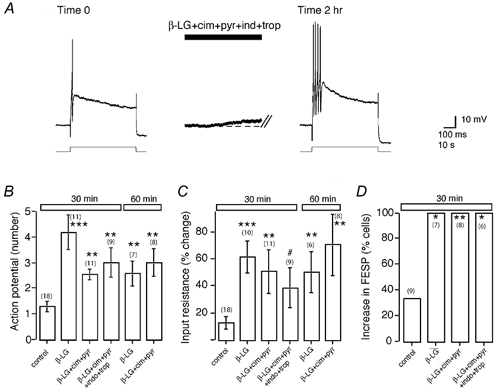
Mast cells that are in close proximity to autonomic and enteric nerves release several mediators that cause neuronal hyperexcitability. This study examined whether mast cell tryptase evokes acute and long-term hyperexcitability in submucosal neurons from the guinea-pig ileum by activating proteinase-activated receptor 2 (PAR2) on these neurons. We detected the expression of PAR2 in the submucosal plexus using RT-PCR. Most submucosal neurons displayed PAR2 immunoreactivity, including those colocalizing VIP. Brief (minutes) application of selective PAR2 agonists, including trypsin, the activating peptide SL-NH2 and mast cell tryptase, evoked depolarizations of the submucosal neurons, as measured with intracellular recording techniques. The membrane potential returned to resting values following washout of agonists, but most neurons were hyperexcitable for the duration of recordings (> 30 min-hours) and exhibited an increased input resistance and amplitude of fast EPSPs. Trypsin, in the presence of soybean trypsin inhibitor, and the reverse sequence of the activating peptide (LR-NH2) had no effect on neuronal membrane potential or long-term excitability. Degranulation of mast cells in the presence of antagonists of established excitatory mast cell mediators (histamine, 5-HT, prostaglandins) also caused depolarization, and following washout of antigen, long-term excitation was observed. Mast cell degranulation resulted in the release of proteases, which desensitized neurons to other agonists of PAR2. Our results suggest that proteases from degranulated mast cells cleave PAR2 on submucosal neurons to cause acute and long-term hyperexcitability. This signalling pathway between immune cells and neurons is a previously unrecognized mechanism that could contribute to chronic alterations in visceral function.
Figures
Similar articles
-
Thrombin and mast cell tryptase regulate guinea-pig myenteric neurons through proteinase-activated receptors-1 and -2.J Physiol. 1999 Jun 15;517 ( Pt 3)(Pt 3):741-56. doi: 10.1111/j.1469-7793.1999.0741s.x. J Physiol. 1999. PMID: 10358115 Free PMC article.
-
Restricted ability of human mast cell tryptase to activate proteinase-activated receptor-2 in rat aorta.Can J Physiol Pharmacol. 2002 Oct;80(10):987-92. doi: 10.1139/y02-125. Can J Physiol Pharmacol. 2002. PMID: 12450065
-
Activity of protease-activated receptors in the human submucous plexus.Gastroenterology. 2011 Dec;141(6):2088-2097.e1. doi: 10.1053/j.gastro.2011.08.034. Epub 2011 Aug 27. Gastroenterology. 2011. PMID: 21875497
-
Proteinase-activated receptors: a growing family of heptahelical receptors for thrombin, trypsin and tryptase.Biochem Soc Trans. 1999 Feb;27(2):246-54. doi: 10.1042/bst0270246. Biochem Soc Trans. 1999. PMID: 10093742 Review. No abstract available.
-
Proteinase-activated receptor-2: physiological and pathophysiological roles.Curr Med Chem Cardiovasc Hematol Agents. 2003 Mar;1(1):61-72. doi: 10.2174/1568016033356715. Curr Med Chem Cardiovasc Hematol Agents. 2003. PMID: 15317291 Review.
Cited by
-
Protease-activated receptor 2 sensitizes the capsaicin receptor transient receptor potential vanilloid receptor 1 to induce hyperalgesia.J Neurosci. 2004 May 5;24(18):4300-12. doi: 10.1523/JNEUROSCI.5679-03.2004. J Neurosci. 2004. PMID: 15128844 Free PMC article.
-
Further Understanding of Neuro-Immune Interactions in Allergy: Implications in Pathophysiology and Role in Disease Progression.J Asthma Allergy. 2022 Sep 10;15:1273-1291. doi: 10.2147/JAA.S282039. eCollection 2022. J Asthma Allergy. 2022. PMID: 36117919 Free PMC article. Review.
-
Research on Correlation Between Psychological Factors, Mast Cells, and PAR-2 Signal Pathway in Irritable Bowel syndrome.J Inflamm Res. 2021 Apr 14;14:1427-1436. doi: 10.2147/JIR.S300513. eCollection 2021. J Inflamm Res. 2021. PMID: 33883919 Free PMC article.
-
The emerging roles of bacterial proteases in intestinal diseases.Gut Microbes. 2023 Jan-Dec;15(1):2181922. doi: 10.1080/19490976.2023.2181922. Gut Microbes. 2023. PMID: 36843008 Free PMC article. Review.
-
Aluminum Ingestion Promotes Colorectal Hypersensitivity in Rodents.Cell Mol Gastroenterol Hepatol. 2018 Sep 20;7(1):185-196. doi: 10.1016/j.jcmgh.2018.09.012. eCollection 2019. Cell Mol Gastroenterol Hepatol. 2018. PMID: 30534582 Free PMC article.
References
-
- Alex G, Furness JB. Sustained post-synaptic excitation in submucosal neurons. Gastroenterology. 2002;122:A38.
-
- Atwood L, James C, Morris GP, Vanner S. Cellular pathways of mast cell- and capsaicin-sensitive nerve-evoked ileal submucosal arteriolar dilations. Am J Physiol. 1998;275:G1063–1072. - PubMed
-
- Barbara G, Cottrell G, Grady E, Cremon C, Di Giogio R, Stanghellini V, Corinaldesi R, Andrade-Gordon P, Bunnett NW. Expression and release of mast cell tryptase in irritable bowel syndrome (IBS) Gastroenterology. 2002;122:A276.
-
- Bauer O, Razin E. Mast cell-nerve interactions. News Physiol Sci. 2000;15:213–218. - PubMed
-
- Bertrand PP, Thomas EA, Kunze WA, Bornstein JC. A simple mathematical model of second-messenger mediated slow excitatory postsynaptic potentials. J Comput Neurosci. 2000;8:127–142. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources