

Modulation of inhibitory autapses and synapses on rat CA1 interneurones by GABA(A) receptor ligands
- PMID: 12562998
- PMCID: PMC2342589
- DOI: 10.1113/jphysiol.2002.035121
Modulation of inhibitory autapses and synapses on rat CA1 interneurones by GABA(A) receptor ligands
Abstract
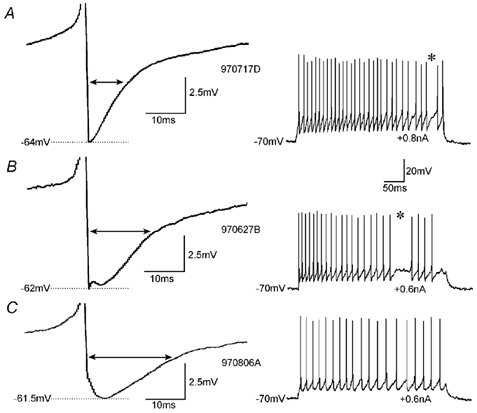
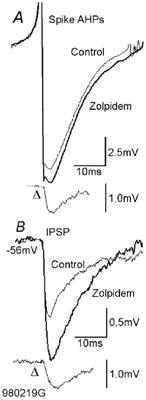
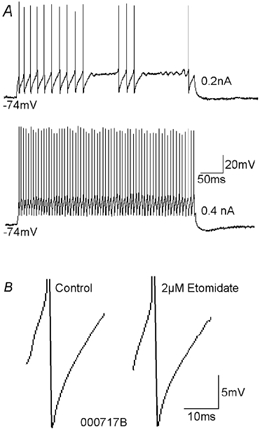
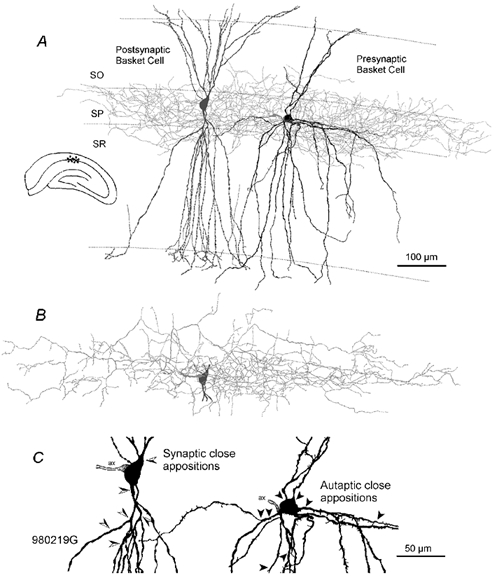
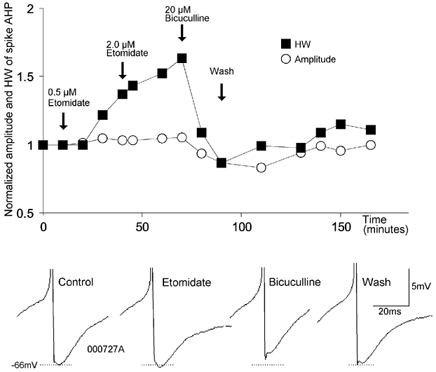
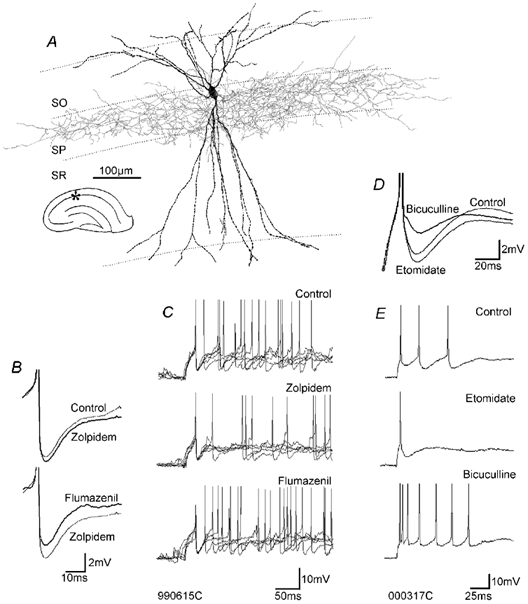
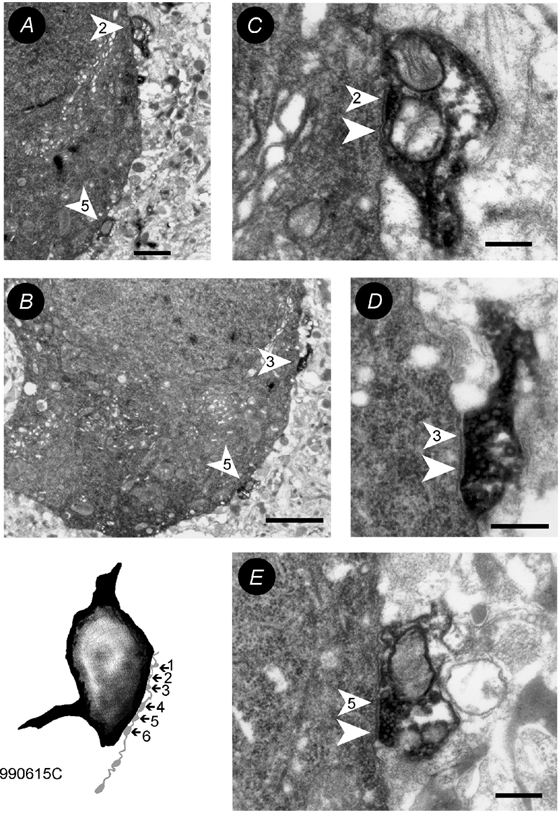
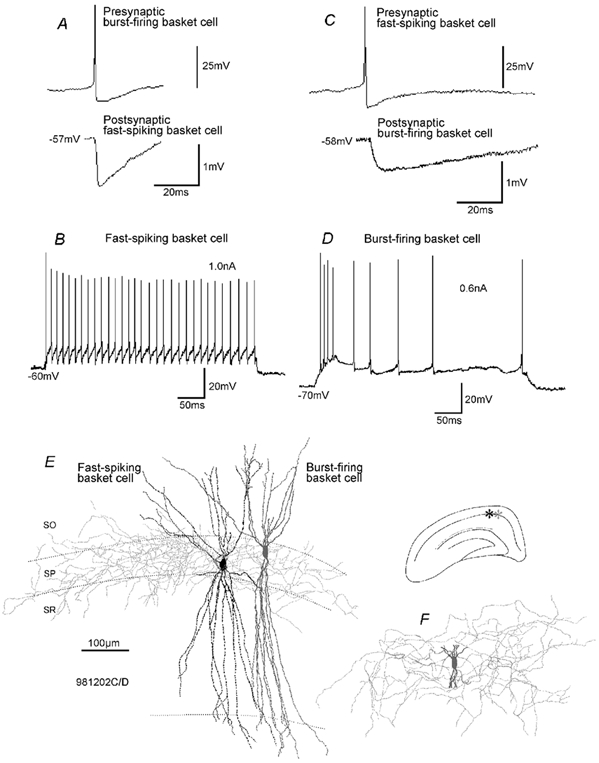
To determine whether autaptic inhibition plays a functional role in the adult hippocampus, the action potential afterhyperpolarisations (spike AHPs) of CA1 interneurones were investigated in 25 basket, three bistratified and eight axo-axonic cells. The spike AHPs showed two minima in all regular-spiking (5), burst-firing (3) and in many fast-spiking cells (17:28). The fast component had a time-to-peak (TTP) of 1.2 +/- 0.5 ms, the slower TTP was very variable (range of 3.3-103 ms). The AHP width at half-amplitude (HW) was 12.5 +/- 5.7 ms in fast-spiking, 29.3 +/- 18 ms in regular-spiking and 99.7 +/- 42 ms in burst-firing cells. Axo-axonic cells never establish autapses, and the fast-spiking variety showed narrow (HW: 3.9 +/- 0.7 ms) spike AHPs with only one AHP minimum (TTP: 0.9 +/- 0.1 ms). When challenged with GABA(A) receptor modulators, spike AHPs in basket and bistratified cells were enhanced by zolpidem (HW by 18.4 +/- 6.2 % in 10:15 cells tested), diazepam (45.2 +/- 0.5 %, 6:7), etomidate (43.9 +/- 36 %, 6:8) and pentobarbitone sodium (41 %, 1:1), and were depressed by bicuculline (-41 +/- 5.7 %, 5:8) and picrotoxin (-54 %, 1:1), and the enhancement produced by zolpidem was reduced by flumazenil (-31 +/- 13 %, relative to the AHP HW during exposure to zolpidem, 3:4). Neuronal excitability was modulated in parallel. The spike AHPs of three axo-axonic cells tested showed no sensitivity to etomidate, pentobarbitone or diazepam. Interneurone-to-interneurone inhibitory postsynaptic potentials (IPSPs), studied with dual intracellular recordings, had time courses resembling those of the spike AHPs. The IPSP HW was 13.4 +/- 2.8 ms in fast-spiking (n = 16) and 28.7 +/- 5.8 ms in regular-spiking/burst-firing cells (n = 6), and the benzodiazepine1-selective modulator zolpidem strongly enhanced these IPSPs (45 +/- 28 %, n = 5). Interneurones with spike AHPs affected by the GABA(A) receptor ligands exhibited 3.8 +/- 1.9 close autaptic appositions. In three basket cells studied at the ultrastructural level 6 of 6, 1 of 2 and 1 of 2 close appositions were confirmed as autapses. Therefore, in the hippocampus autaptic connections contribute to spike AHPs in many interneurones. These autapses influence neuronal firing and responses to GABA(A) receptor ligands.
Figures
References
-
- Ali AB, Bannister AP, Thomson AM. IPSPs elicited in CA1 pyramidal cells by putative basket cells in slices of adult rat hippocampus. Eur J Neurosci. 1999;11:1741–1753. - PubMed
-
- Aoki T, Baraban SC. Properties of a calcium-activated K+ current on interneurons in the developing rat hippocampus. J Neurophysiol. 2000;83:3453–3461. - PubMed
-
- Buhl EH, Han Z-S, Lörinczi Z, Stezhka VV, Karnup SV, Somogyi P. Physiological properties of anatomically identified axo-axonic cells in the rat hippocampus. J Neurophysiol. 1994;71:1289–1307. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous