

Extrathymic T cell lymphopoiesis: ontogeny and contribution to gut intraepithelial lymphocytes in athymic and euthymic mice
- PMID: 12566417
- PMCID: PMC2193840
- DOI: 10.1084/jem.20021639
Extrathymic T cell lymphopoiesis: ontogeny and contribution to gut intraepithelial lymphocytes in athymic and euthymic mice
Abstract
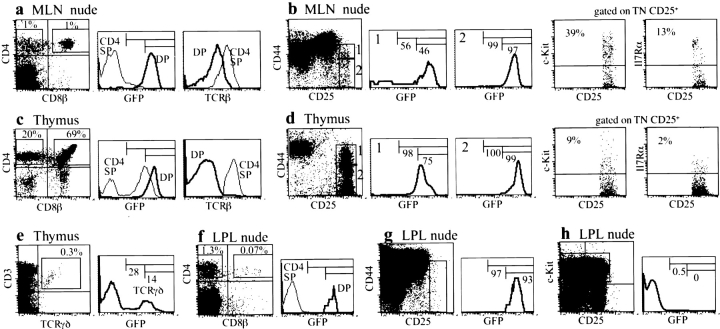
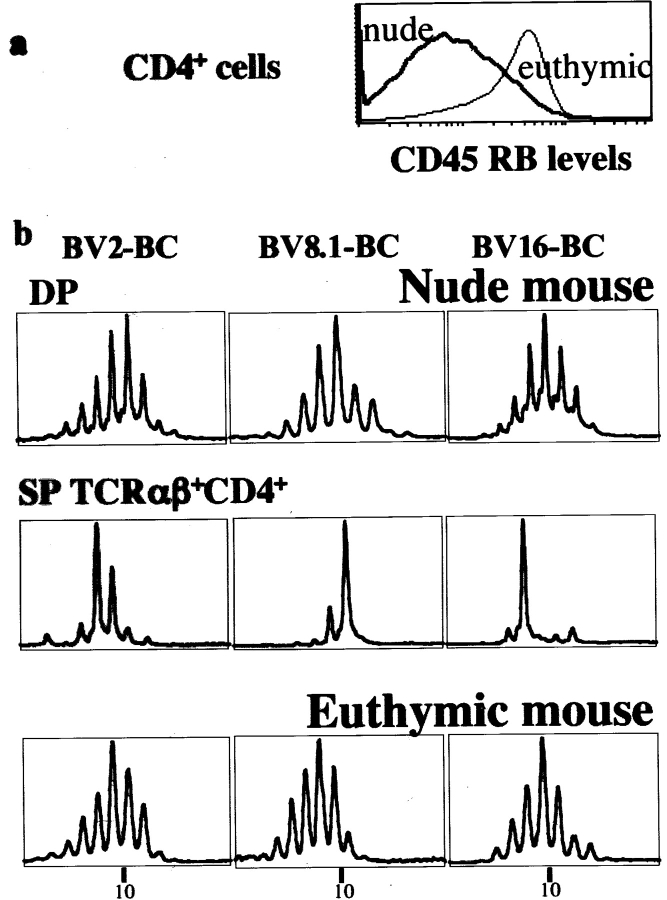
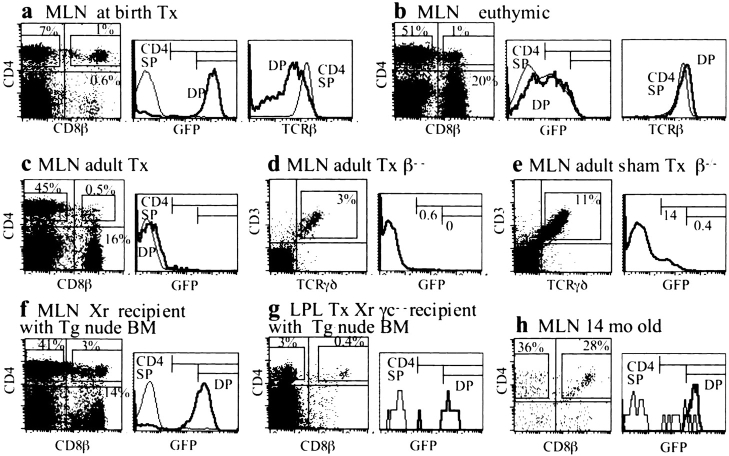
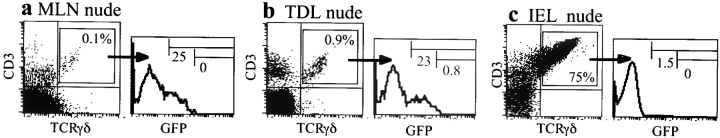
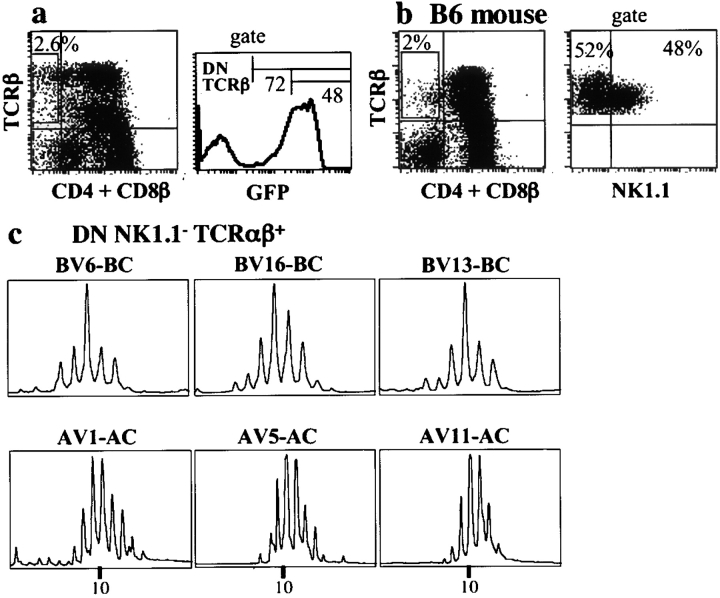
In the absence of thymopoiesis, T lymphocytes are nevertheless present, mainly in the gut epithelium. Ontogeny of the extrathymic pathway and the extent of its involvement in euthymic mice are controversial. These questions have been addressed by assessing the expression of recombinase activating gene (RAG) through the use of green fluorescent protein RAG2 transgenic mouse models. In athymic mice, T lymphopoiesis occurs mainly in the mesenteric lymph node and less in the Peyer's patches. Ontogenic steps of this lymphopoiesis resemble those of thymopoiesis, but with an apparent bias toward gamma delta T cell production and with a paucity of oligoclonal alpha beta T cells possibly resulting from a deficit in positive selection. Whether in athymic or euthymic mice, neither T intraepithelial lymphocytes (IEL) nor cryptopatch cells (reported to contain precursors of IEL) displayed fluorescence indicating recent RAG protein synthesis. Newly made T cells migrate from the mesenteric node into the thoracic duct lymph to reach the gut mucosa. In euthymic mice, this extrathymic pathway is totally repressed, except in conditions of severe lymphocytic depletion. Thus, in normal animals, all gut T IEL, including CD8 alpha alpha(+) cells, are of thymic origin, CD8 alpha alpha(+) TCR alpha beta(+) IEL being the likely progeny of double negative NK1-1(-) thymocytes, which show polyclonal V alpha and V beta repertoires.
Figures

References
-
- Park, S.H., D. Guy-Grand, F.A. Lemonnier, C.R. Wang, A. Bendelac, and B. Jabri. 1999. Selection and expansion of CD8αα+ T cell receptor αβ+ intestinal intraepithelial lymphocytes in the absence of both classical major histocompatibility complex class I and nonclassical CD1 molecules. J. Exp. Med. 190:885–890. - PMC - PubMed
-
- Gapin, L., H. Cheroutre, and M. Kronenberg. 1999. TCR αβ+ CD8 αα+ T cells are found in intestinal intraepithelial lymphocytes of mice that lack classical MHC class I molecules. J. Immunol. 163:4100–4104. - PubMed
-
- Poussier, P., and M. Julius. 1994. Thymus independent T cell development and selection in the intestinal epithelium. Annu. Rev. Immunol. 12:521–553. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
