

The poxvirus protein A52R targets Toll-like receptor signaling complexes to suppress host defense
- PMID: 12566418
- PMCID: PMC2193841
- DOI: 10.1084/jem.20021652
The poxvirus protein A52R targets Toll-like receptor signaling complexes to suppress host defense
Abstract
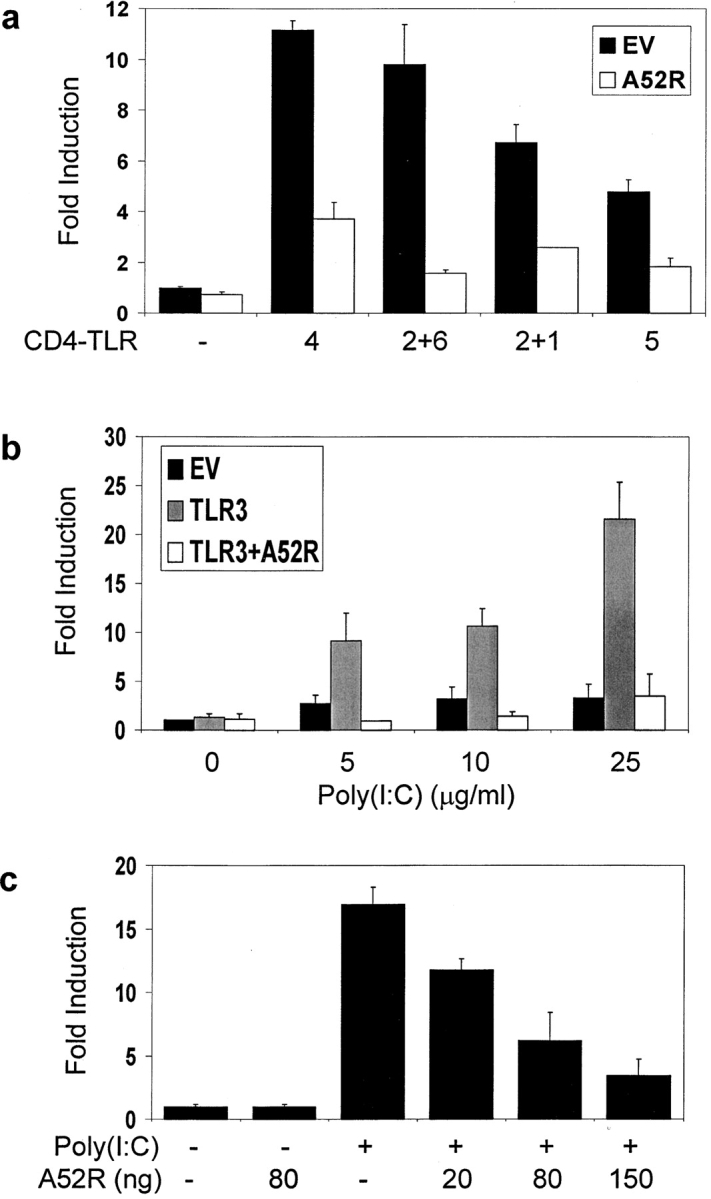
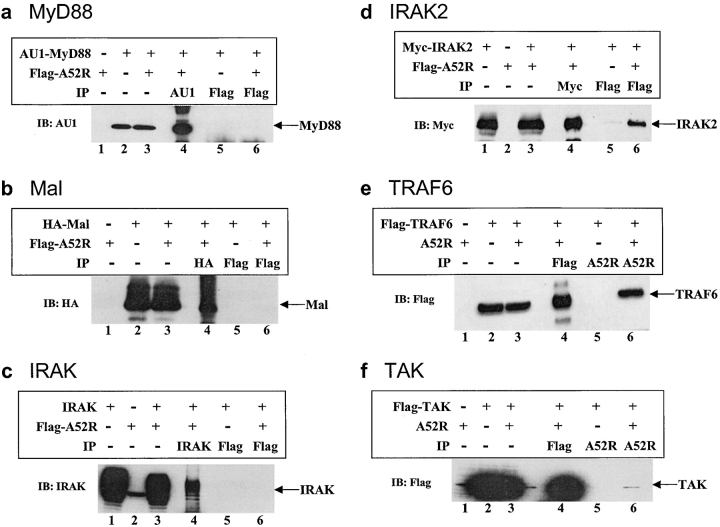
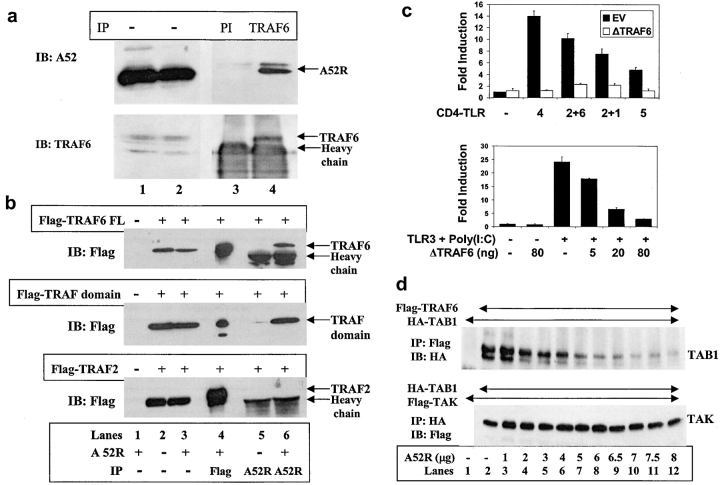
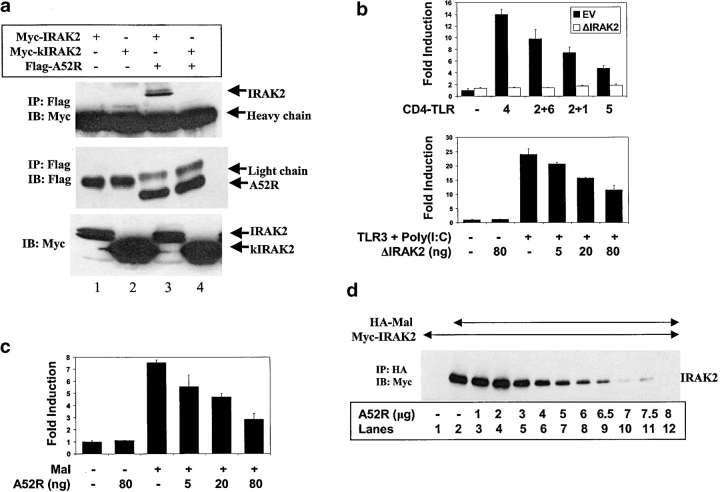
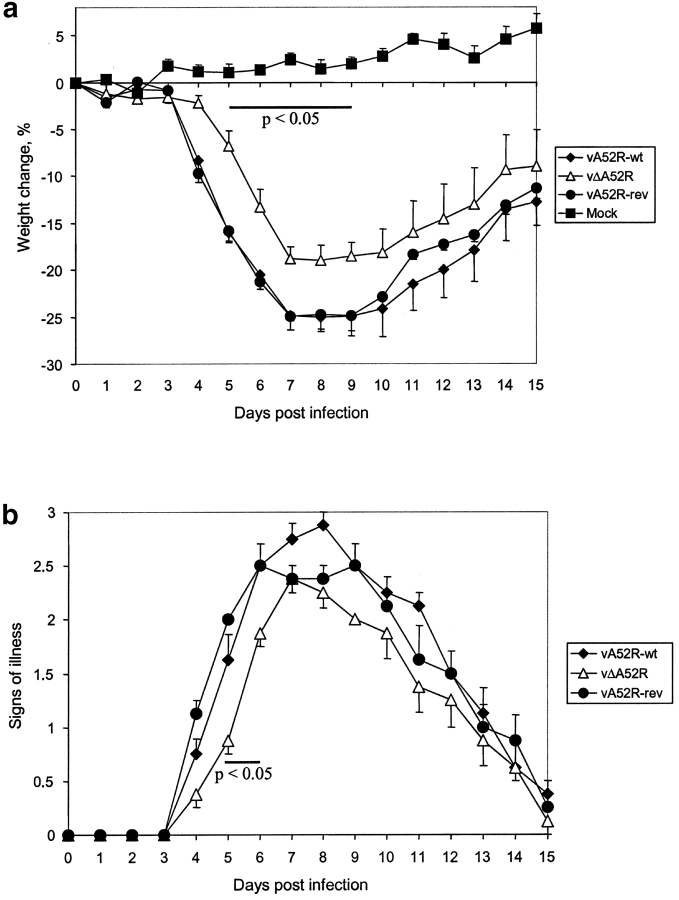
Toll-like receptors (TLRs) are crucial in the innate immune response to pathogens, in that they recognize and respond to pathogen associated molecular patterns, which leads to activation of intracellular signaling pathways and altered gene expression. Vaccinia virus (VV), the poxvirus used to vaccinate against smallpox, encodes proteins that antagonize important components of host antiviral defense. Here we show that the VV protein A52R blocks the activation of the transcription factor nuclear factor kappa B (NF-kappa B) by multiple TLRs, including TLR3, a recently identified receptor for viral RNA. A52R associates with both interleukin 1 receptor-associated kinase 2 (IRAK2) and tumor necrosis factor receptor-associated factor 6 (TRAF6), two key proteins important in TLR signal transduction. Further, A52R could disrupt signaling complexes containing these proteins. A virus deletion mutant lacking the A52R gene was attenuated compared with wild-type and revertant controls in a murine intranasal model of infection. This study reveals a novel mechanism used by VV to suppress the host immunity. We demonstrate viral disabling of TLRs, providing further evidence for an important role for this family of receptors in the antiviral response.
Figures
References
-
- Akira, S., K. Takeda, and T. Kaisho. 2001. Toll-like receptors: critical proteins linking innate and acquired immunity. Nat. Immunol. 2:675–680. - PubMed
-
- Bowie, A., and L.A.J. O'Neill. 2000. The interleukin-1 receptor/Toll-like receptor superfamily: signal generators for pro-inflammatory interleukins and microbial products. J. Leukoc. Biol. 67:508–514. - PubMed
-
- Poltorak, A., X. He, I. Smirnova, M.-Y. Liu, C. Van Huffel, X. Du, D. Birdwell, E. Alejos, M. Silva, C. Galanos, et al. 1998. Defective LPS signalling in C3H/HeJ and C57BL/10ScCr mice: mutations in the Tlr4 gene. Science. 282:2085–2088. - PubMed
-
- Hayashi, F., K.D. Smith, A. Ozinsky, T.R. Hawn, E.C. Yi, D.R. Goodlett, J.K. Eng, S. Akira, D.M. Underhill, and A. Aderem. 2001. The innate immune response to bacterial flagellin is mediated by Toll-like receptor 5. Nature. 410:1099–1103. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
