

STAT3 deletion during hematopoiesis causes Crohn's disease-like pathogenesis and lethality: a critical role of STAT3 in innate immunity
- PMID: 12571365
- PMCID: PMC149927
- DOI: 10.1073/pnas.0237137100
STAT3 deletion during hematopoiesis causes Crohn's disease-like pathogenesis and lethality: a critical role of STAT3 in innate immunity
Abstract
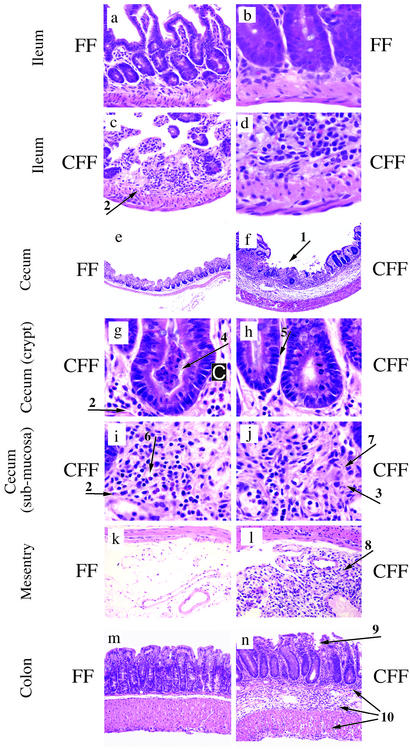
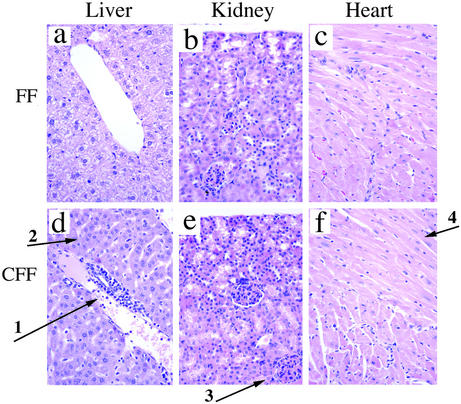
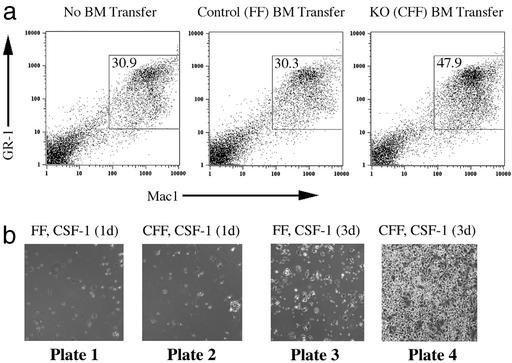
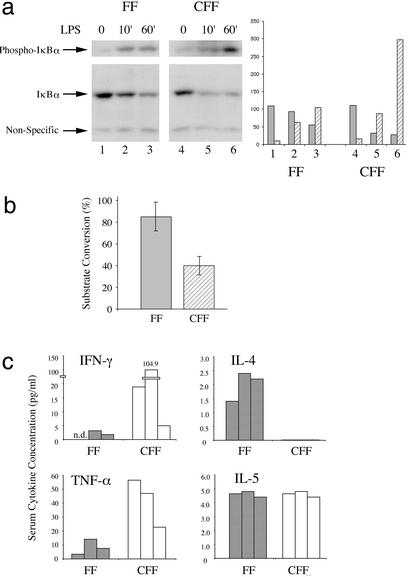
Signal transducer and activator of transcription 3 (STAT3) is a key transcriptional mediator for many cytokines and is essential for normal embryonic development. We have generated a unique strain of mice with tissue-specific disruption of STAT3 in bone marrow cells during hematopoiesis. This specific STAT3 deletion causes death of these mice within 4-6 weeks after birth with Crohn's disease-like pathogenesis in both the small and large intestine, including segmental inflammatory cell infiltration, ulceration, bowel wall thickening, and granuloma formation. Deletion of STAT3 causes significantly increased cell autonomous proliferation of cells of the myeloid lineage, both in vivo and in vitro. Most importantly, Stat3 deletion during hematopoiesis causes overly pseudoactivated innate immune responses. Although inflammatory cytokines, including tumor necrosis factor alpha and IFN-gamma, are overly produced in these mice, the NAPDH oxidase activity, which is involved in antimicrobial and innate immune responses, is inhibited. The signaling responses to lipopolysaccharide are changed in the absence of STAT3, leading to enhanced NF-kappa B activation. Our results suggest a model in which STAT3 has critical roles in the development and regulation of innate immunity, and deletion of STAT3 during hematopoiesis results in abnormalities in myeloid cells and causes Crohn's disease-like pathogenesis.
Figures

References
-
- Medzhitov R, Janeway C A., Jr Cell. 1997;91:295–298. - PubMed
-
- Strober W, Fuss I J, Blumberg R S. Annu Rev Immunol. 2002;20:495–549. - PubMed
-
- Hugot J P, Zouali H, Lesage S, Thomas G. Int J Colorectal Dis. 1999;14:2–9. - PubMed
-
- Laroux F S, Pavlick K P, Wolf R E, Grisham M B. News Physiol Sci. 2001;16:272–277. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
