

Transient and progressive electrophysiological alterations in the corticostriatal pathway in a mouse model of Huntington's disease
- PMID: 12574425
- PMCID: PMC6741903
- DOI: 10.1523/JNEUROSCI.23-03-00961.2003
Transient and progressive electrophysiological alterations in the corticostriatal pathway in a mouse model of Huntington's disease
Abstract
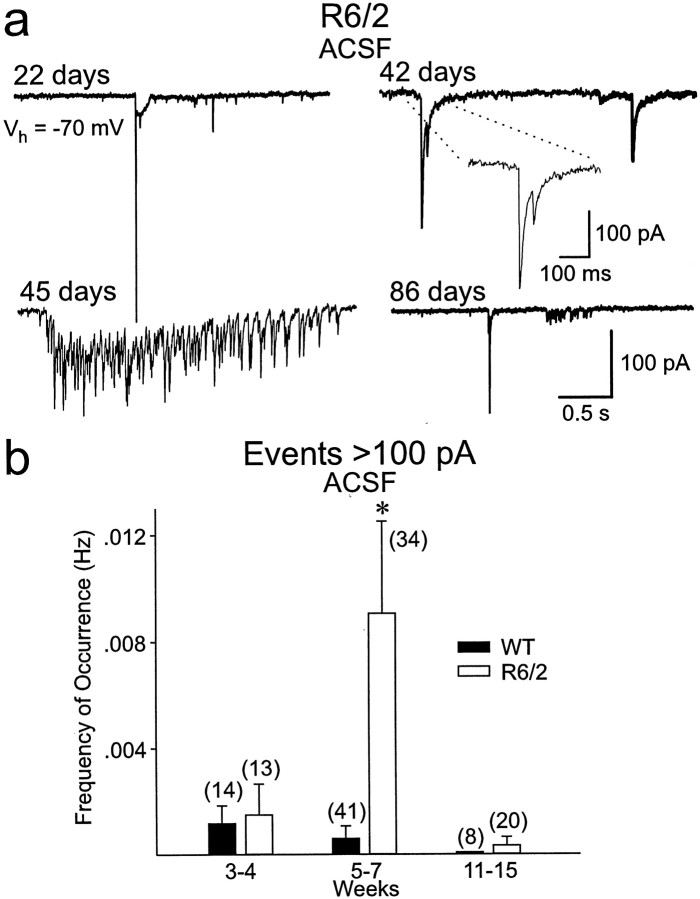
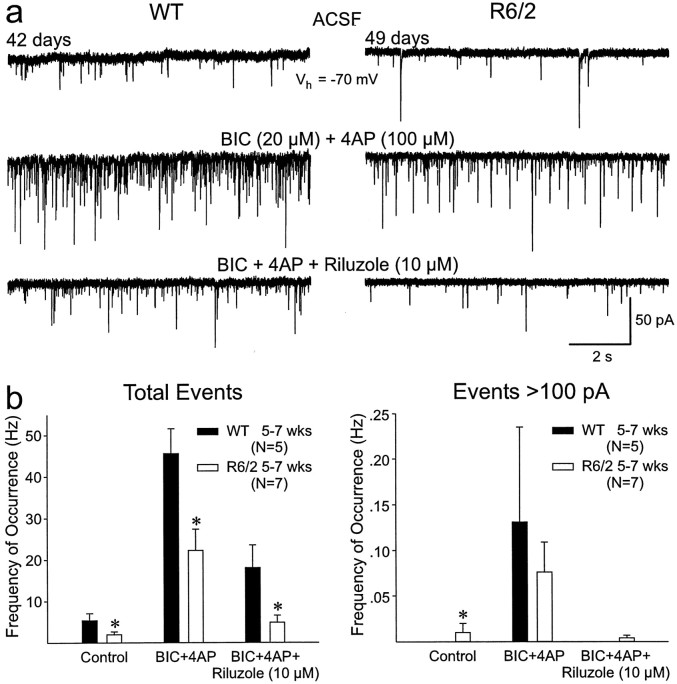
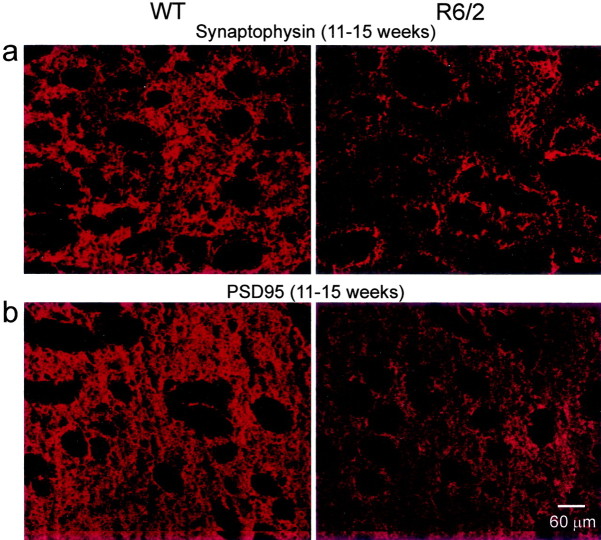
Alterations in the corticostriatal pathway may precede symptomatology and striatal cell death in Huntington's disease (HD) patients. Here we examined spontaneous EPSCs in striatal medium-sized spiny neurons in slices from a mouse model of HD (R6/2). Spontaneous EPSC frequency was similar in young (3-4 weeks) transgenics and controls but decreased significantly in transgenics when overt behavioral symptoms began (5-7 weeks) and was most pronounced in severely impaired transgenics (11-15 weeks). These differences were maintained after bicuculline or tetrodotoxin, indicating they were specific to glutamatergic input and likely presynaptic in origin. Decreases in presynaptic and postsynaptic protein markers, synaptophysin and postsynaptic density-95, occurred in 11-15 week R6/2 mice, supporting the electrophysiological results. Furthermore, isolated, large-amplitude synaptic events (>100 pA) occurred more frequently in transgenic animals, particularly at 5-7 weeks, suggesting additional dysregulation of cortical inputs. Large events were blocked by tetrodotoxin, indicating a possible cortical origin. Addition of bicuculline and 4-aminopyridine facilitated the occurrence of large events. Riluzole, a compound that decreases glutamate release, reduced these events. Together, these observations indicate that both progressive and transient alterations occur along the corticostriatal pathway in experimental HD. These alterations are likely to contribute to the selective vulnerability of striatal medium-sized spiny neurons.
Figures
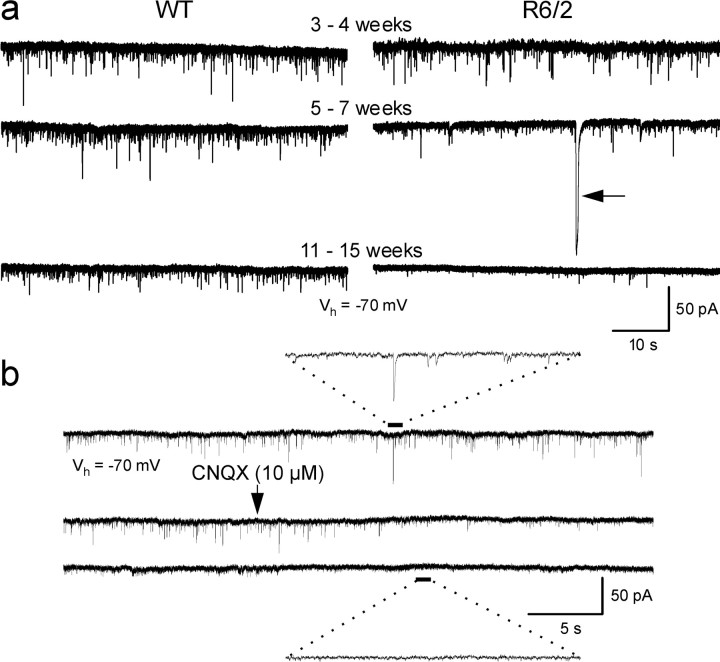
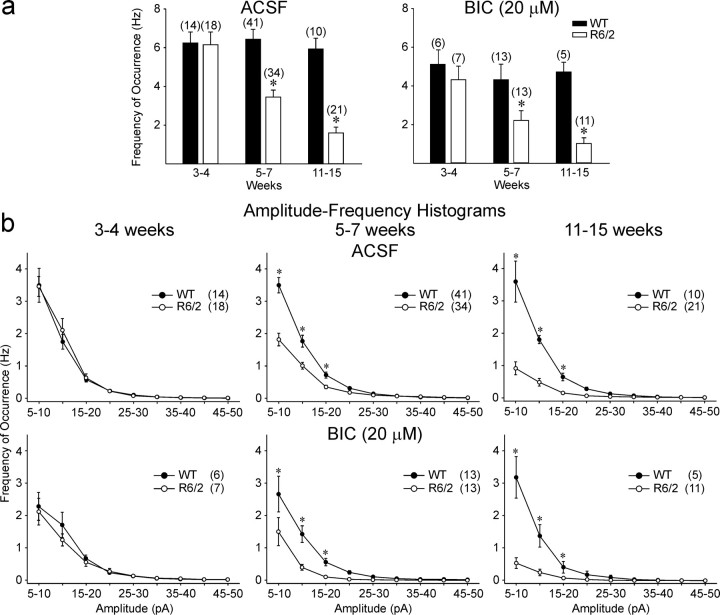
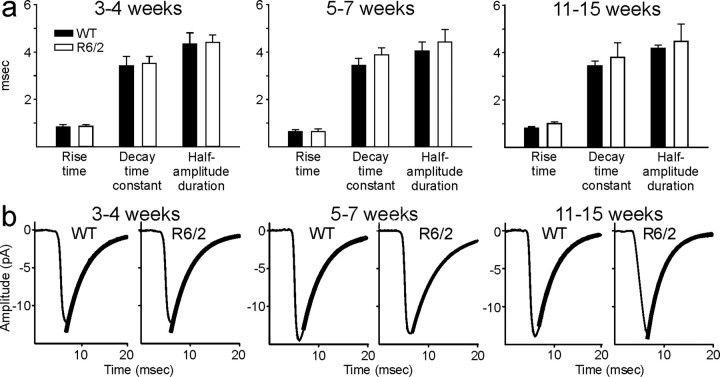
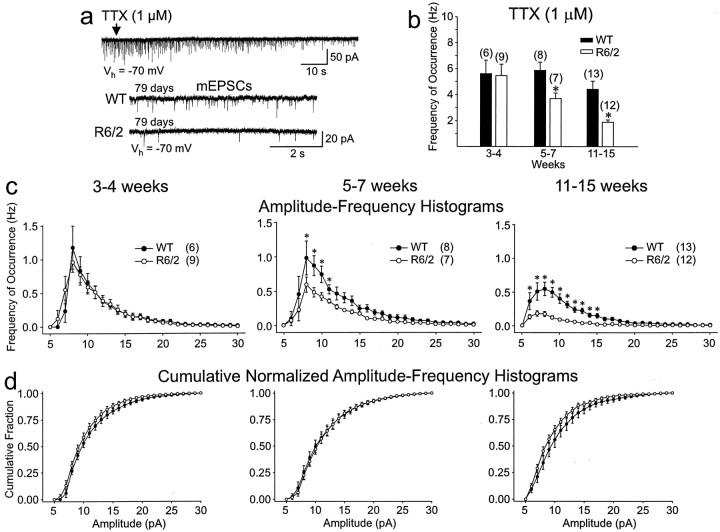
References
-
- Ariano MA, Aronin N, DiFiglia M, Tagle DA, Sibley DR, Leavitt BR, Hayden MR, Levine MS. Striatal neurochemical changes in transgenic models of Huntington's disease. J Neurosci Res. 2002;68:716–729. - PubMed
-
- Behrens PF, Franz P, Woodman B, Lindenberg KS, Landwehrmeyer GB. Impaired glutamate transport and glutamate-glutamine cycling: downstream effects of the Huntington mutation. Brain. 2002;125:1908–1922. - PubMed
-
- Biziere K, Coyle JT. Effects of cortical ablation on the neurotoxicity and receptor binding of kainic acid in striatum. J Neurosci Res. 1979;4:383–398. - PubMed
-
- Calabresi P, Picconi B, Saulle E, Centonze D, Hainsworth AH, Bernardi G. Is pharmacological neuroprotection dependent on reduced glutamate release? Stroke. 2000;31:766–772. - PubMed
-
- Calabresi P, Gubellini P, Picconi B, Centonze D, Pisani A, Bonsi P, Greengard P, Hipskind RA, Borrelli E, Bernardi G. Inhibition of mitochondrial complex II induces a long-term potentiation of NMDA-mediated synaptic excitation in the striatum requiring endogenous dopamine. J Neurosci. 2001;21:5110–5120. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases